Rocky subtidal habitatには、海底のすべての硬い基質領域が含まれています。 地質起源の基質成分には,CMECの非固結鉱物基質クラスの玉石とボルダー,岩石基質クラスの岩盤とメガクラストが含まれる。 人類起源のハード基板もここにあります。 人為的なサンゴ礁には、硬くて永続的な材料が意図的にまたは偶発的に人間によって配置された領域が含まれます。 例としては、多くの湾の入り口にある岩の突堤、難破船、再生可能エネルギープロジェクトのための固定システム、水中ケーブルやパイプラインの未埋 岩質の亜潮域は、しばしばサンゴ礁、岩のサンゴ礁、岩の銀行、ピナクルズ、または”ハードボトム”と呼ばれています。”自然と人為的なコンポーネントの両方を含む岩の潮下の生息地は、近海と沖合の海洋サブシステムの潮下のゾーン内にあるとしてCMECSによって特徴付けら ほとんどの地域は空気にさらされることはありませんが、CMECSサブ潮位の定義には、潮位が平均低水位(MLLW)レベルを下回ったときに毎月断続的に露出してい 岩質の亜潮の生息地は、ニアショアサブシステムとオフショアサブシステムの両方で発見され、違いのいくつかは以下で議論されています。
いくつかの岩質の亜潮域は、岬、崖、または岩質の潮間帯の生息地などの海岸線の岩の特徴の延長であり、他のものは柔らかい底質の生息地に囲まれた岩の孤立した領域として存在する。 岩礁は地形が多様であり、周囲の海底の上にほとんど来ないものもあれば、海底から何メートルも上昇したり、海面の上に伸びて領海に島を形成したりするものもある。 オレゴン州の沖には1,800以上の島があり、その基地は岩場の亜潮の生息地を形成しています。
物理的環境
岩質下の生息地の物理的特性は、海岸への近さ、水深、地元の海底地質、侵食力、生物学的影響を反映しています。 多くの岩の亜潮域の地質は、しばしばオレゴン州の岩の岬で一般的な侵食抵抗性玄武岩または変成岩からなる、隣接する地形の地質学を模倣して 地質学的には、連続した氷河期と地質学的な力が大規模な海面変化を引き起こしたため、水中の岩石の特徴は隆起し、曲がり、変形し、海洋と陸の侵食力に交互に曝されてきた。 これらの力は、平らな岩のベンチ、スタック、ギザギザの尾根、壊れたボルダーフィールド、豊富な生活に避難所と基板を提供する亀裂や割れ目の膨大な数を
海洋学的プロセスと特徴は、岩場の潮下環境に強く影響します。 潮下のサンゴ礁は、波の作用、水中の流れ、および水の物理的および化学的性質にさらされています。 これらの要因は、サンゴ礁の生物群集に影響を与えます。 一般的に、沿岸のサンゴ礁は沖合のサンゴ礁よりも波の作用にさらされており、波の作用は夏よりも冬にははるかに強い。 波の作用は、非常に浅いサンゴ礁に住むことができる生物の種類を決定する上で重要な要因です。 海流は、場所、年の時間、および潮汐サイクルによって大きく異なります。 海流は、直接侵食、砂の洗掘やサンゴ礁の埋葬、プランクトンや幼虫を含むサンゴ礁への生物の移動など、さまざまな方法でサンゴ礁に影響を与えます。 湧昇、季節的な現在の方向シフト、海洋循環のシフト、水温変動、局所的および全球的な気象パターン、海洋酸性化、および生物学的プロセスなどの海洋環境の大規模または長期的な変動は、岩場下の生息地における水の周囲の化学的および物理的組成を決定するために組み合わされる。 CMECS水柱コンポーネントは、そこに住んでいる生物学的コミュニティを形成する上で重要である岩礁を囲むと上に水の重要な特徴を記述するために使
30mの深さの輪郭は、CMECSによってnearshoreサブシステムとoffshoreサブシステムの境界として定義されます。 近海の岩礁は、いくつかの重要な物理的特性において沖合の岩礁とは異なる。 光の浸透は、沿岸のサンゴ礁で藻類の生活を支えるのに十分ですが、沖合のサンゴ礁では藻類の成長がはるかに少なくなります。 例えば、ケルプは海岸近くの岩場にしか見られない。 波の作用、流れ、および嵐は、より深いサンゴ礁よりも沿岸のサンゴ礁でより高いエネルギー環境を作り出します。 より高いエネルギー環境に適応した生物は、沿岸地域でより一般的である。 いくつかのサンゴ礁では、強い流れが砂で岩を磨き、季節的に埋めたり、露出させたりすることがあり、それらの岩礁下の環境を利用できる生物の種類
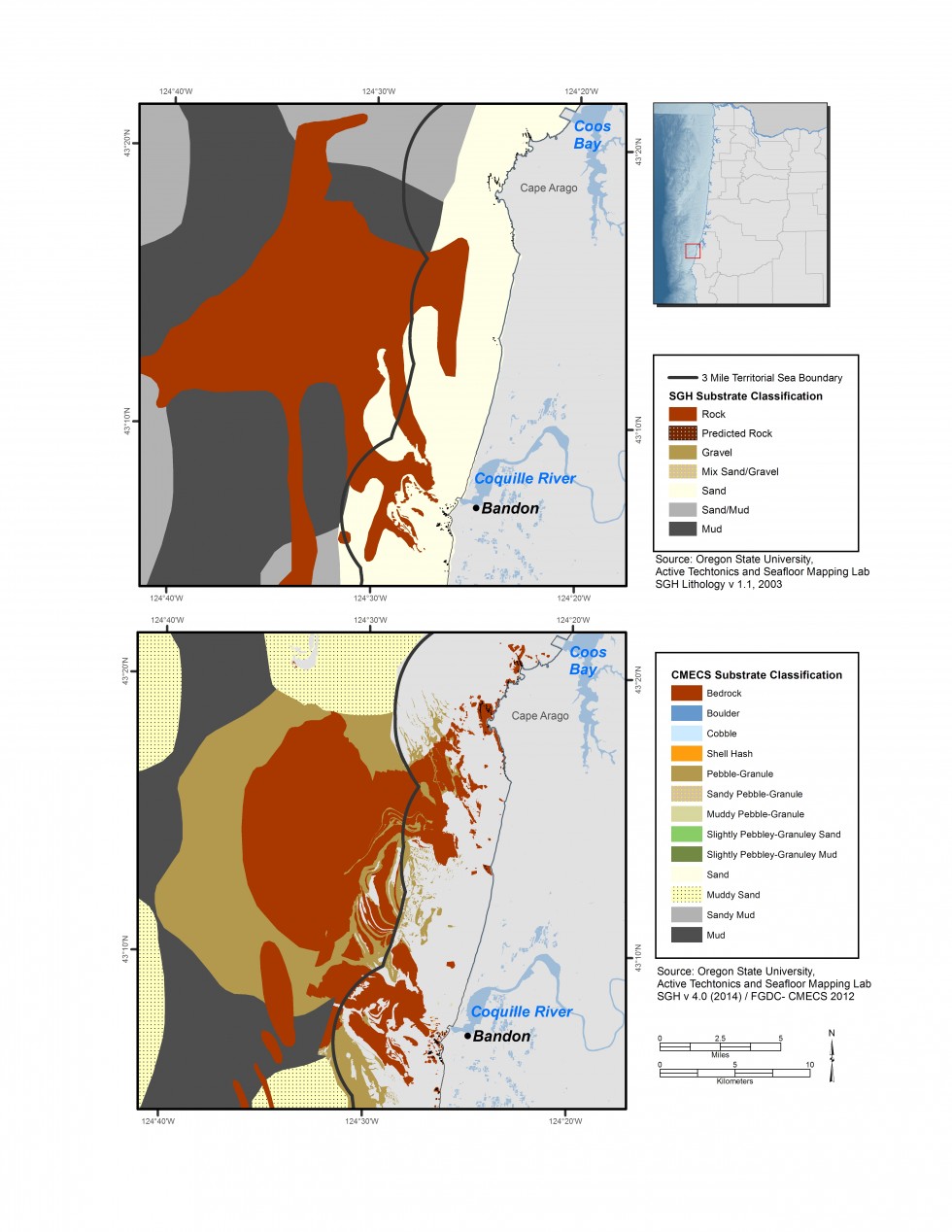
新しい生息地マップの詳細な違いは、2006年版のニアショア戦略で利用可能なものと比較して顕著です。 CMECSの基質の構成システムの使用はまたずっとより多くの細部を提供する。 その良い例は、アラゴ岬沖のエリアです(図6.7)大規模で相違が非常に明白であるところで。
生物学的特性
潮下の岩礁は、豊富で多様な生物群集で知られています。 岩礁内および岩礁間の地形、基質特性、および深さの多様性は、しばしば比較的小さな地理的領域内で、大量のマイクロハビタットを生成する。 これは、順番にこれらの異なるmicrohabitatsでの生活に適応した種の多様性を提供します。 昆布や付着した無脊椎動物などの生息地を形成する生物は、サンゴ礁種によって使用される追加のマイクロハビタットを提供する。
ほとんどの沿岸の岩礁は、豊富な藻類、無脊椎動物、魚、鳥、海洋哺乳類のコミュニティを持っています。 水深、光の浸透、波のエネルギー、および他の物理的および生物学的プロセスに応じて、藻類および大藻類は、沿岸サブシステム内の他の種に広範または散発的な覆いおよび食物を提供することができる。 藻およびmacroalgaeは石の表面の近くで育つ覆いの形態、フィートの厚いまたは多くまで密な層を作成できる泥炭の形態、加えられた表面下の生息地の構造を提 深海の沖合の岩礁には昆布の森はありません。 フリー水泳(nektonic)、漂流(planktonic)、および接続された無脊椎動物は、沿岸および沖合いの岩の亜潮の生息地の両方で一般的です。
多くの戦略、監視リスト、およびその他の一般的に関連する種は、岩場の亜種の生息地に生息しています(表6.3、付録EおよびF)。 黒、青、中国、ディーコン、銅とキルバック岩魚、ウルフウナギ、パイルとストリップパーチ、lingcod、cabezonとgreenlingsなどの魚は、小さなsculpins、gunnels、密猟者、blenniesなどの多種多様な岩のsubtidalの生息地に関連付けられています。 ダイビングの海鳥および海洋ほ乳類は岩が多いsubtidal区域で広く飼料する。 さまざまなフィルターまたは懸濁液供給無脊椎動物は、スポンジ、イソギンチャク、フジツボ、コケ動物、ハイドロゾアン、tunicates、および冷水サンゴなどの硬い基材に付着します。 モバイル無脊椎動物は、同様にここにたくさんあります。 赤と紫のウニ、赤と平らなアワビは岩に付着した藻類を食べる。 黄土色、ヒマワリおよび他の海の星はカニ、エビ、壊れやすい星、nudibranchs、キトン、およびみみずがそうであるようにsubtidal岩が多い生息地で餌を与えます。
岩質の亜潮に見られる生産者と消費者の多様性は、複雑な食物網と生物間の相互依存関係を作り出している。 サンゴ礁は、海流や生物の動きによって周囲の環境にリンクされています。 サンゴ礁の地形構造は、しばしば流れを遅くし、漂流生物を捕獲する地域社会の能力を高め、大きな昆布ベッドの時折の存在によって強化された効果。 多くの生物はサンゴ礁の内外を移動し、いくつかは大規模な移動であり、他の地域への短い摂食の進出である。 ほとんどの沿岸サンゴ礁の魚は、沿岸と沖合の両方のサンゴ礁を占めていますが、いくつかの種と生活史の段階の深さの好みに違いがあります。
いくつかの魚種は、成長するにつれてより深いサンゴ礁、大陸棚、または他の地域に移動する前に、初期の生活史段階で沿岸の岩礁に依存しています。 逆に、いくつかの魚は、成体として岩の潮下地域に移動する前に、初期の生活史の段階のための河口や岩の潮間帯の生息地に依存しています。 例えば、ケルプgreenling、cabezon、および草のロックフィッシュは、沿岸のサンゴ礁でより一般的になる傾向があります。 カナリアとイエローアイロックフィッシュは、成長するにつれて沿岸から沖合のサンゴ礁に移動します。 他の人が訪問者である間、多くの魚種は、そのライフサイクルの一部のためにサンゴ礁に完全に依存しています。 一般的な訪問者には、ニシン、ワカサギ、サメ、ラットフィッシュ、サケが含まれます。
岩質下の生息地との間の生態学的なつながりは、その生物群集とこの生息地タイプに見られる種の多様性を形成するのに役立ちます。 流れはプランクトン性の有機体をもたらし、異種の岩の亜潮の生息地に出入りして漂流幼虫を輸送します。 他の「上流」または「下流」のサンゴ礁に対するサンゴ礁の位置は、サンゴ礁のコミュニティおよび生物の種類、豊富さ、および募集率に劇的な影響を与え 生物の相互関係のこの複雑さは、サンゴ礁への自然または人間の妨害の結果を測定または予測することを困難にする。
追加生物成分:昆布床

昆布のベッドは、オレゴン州の岩質の亜潮の生息地の重要なサブセットです。 CMECSは、ケルプベッドをオレゴン州の岩場下の生息地の生物成分として、より具体的にはキャノピー形成藻類ベッドとして分類しています。 オレゴン州の海岸近くの岩礁の多くに見られる昆布のベッドは、一般的に海底から海面に成長し、昆布の浮遊キャノピーを形成する茶色の大藻類の一種以上の集合体で構成されています。 ケルプベッドはオレゴン州の海岸に沿ってすべて見つけることができますが、アラゴ岬から南の海岸帯には州のケルプベッドの約92%が含まれています(図6.8aと6.8b)。 オレゴン州のほとんどの昆布ベッドはブル昆布(Nereocystis luetkeana)で構成されています。 ケルプベッドは海岸からの視界のために一般的に見えるが、実際にはオレゴン州の水域では比較的希少な生息地であり、沿岸地域の一パーセント未満をカバーしている。
昆布床の存在と属性は、多くの物理的および生物学的変数に依存します。 昆布が存在する可能性がある場所を決定する主な変数には、水深と基質の可用性が含まれます。 オレゴン州の水域では、昆布の床は岩の基質上にのみ形成され、沿岸サブシステムに限定されています。 その深さを超えて、海底の低い光のレベルは、昆布の成長を制限します。 しかし、光と基質だけが制限要因ではなく、適切な深さの範囲にある多くの岩礁は、昆布のベッドをサポートすることはめったにありません。 これらのサンゴ礁の昆布を制限する要因には、サンゴ礁の季節的な砂の埋葬、岩の砂の洗浄、波と嵐のエネルギーへの過度の露出、局所的に高い濁度、栄養素の欠如、昆布の”播種”源へのサンゴ礁の距離、昆布を消費する生物(例えば、ウニ)の豊富さ、および付着のために利用可能な岩石基質のための無脊椎動物および他の藻類との競争が含まれる。
オレゴン州の昆布床は、季節的および年間的な範囲および密度の変化を顕著に示しています。 ブル昆布ベッドは嵐がほとんど、あるいはまったく表面のキャノピーを残して、藻類の多くを取り除く冬の期間に続いて、春と夏に急速に成長します。 昆布床のバイオマスは、その成長に影響を与える物理的および生物学的変数の組み合わせの年次変動のために、毎年10倍以上変化する可能性があ
昆布床は、昆布の一次生産性と昆布床が周囲の環境に及ぼす影響の両方のために、生物学的に豊富な生息地です。 ブル昆布は、世界で最も急速に成長している生物の一つであり、毎年、昆布が死んだ後に直接またはデトリタスとして消費するために利用可能な大規模なバイオマスを提供しています。 ケルプは、そうでなければサンゴ礁には存在しない垂直の生息地構造を提供します。 昆布のベッドはまた水流を遅らせ、波および風のチョップを減らし、漂流幼虫および栄養素を引っ掛けるのを助け、避難所を提供する。
昆布の床とその天蓋は、藻類と付着した無脊椎動物のカバーの豊かな下層を支えることもできます。 オレゴンのサンゴ礁では、密な下層藻類のカバレッジは、約5-10mの水深で支配的な無脊椎動物のカバーに道を譲ります。 厚い昆布カバーは軽い浸透を減らし、下層藻類の密度を制限することができます。 昆布の床とその下にあるサンゴ礁は、魚や無脊椎動物の種の多様な配列をサポートし、ダイビング海鳥や海洋哺乳類のためのカバーと採餌エリアを提供 オレゴン州では、昆布のベッドと非昆布のベッドのサンゴ礁の魚種のミックスは似ています。 昆布のベッドが研究されている世界のほとんどの地域では、昆布のベッドを持つサンゴ礁は、昆布のない同様のサンゴ礁よりもはるかに高い密度の魚 オレゴン州では、これはそうではないようです。 しかし、これを確認するための定量的比較研究は行われていない。
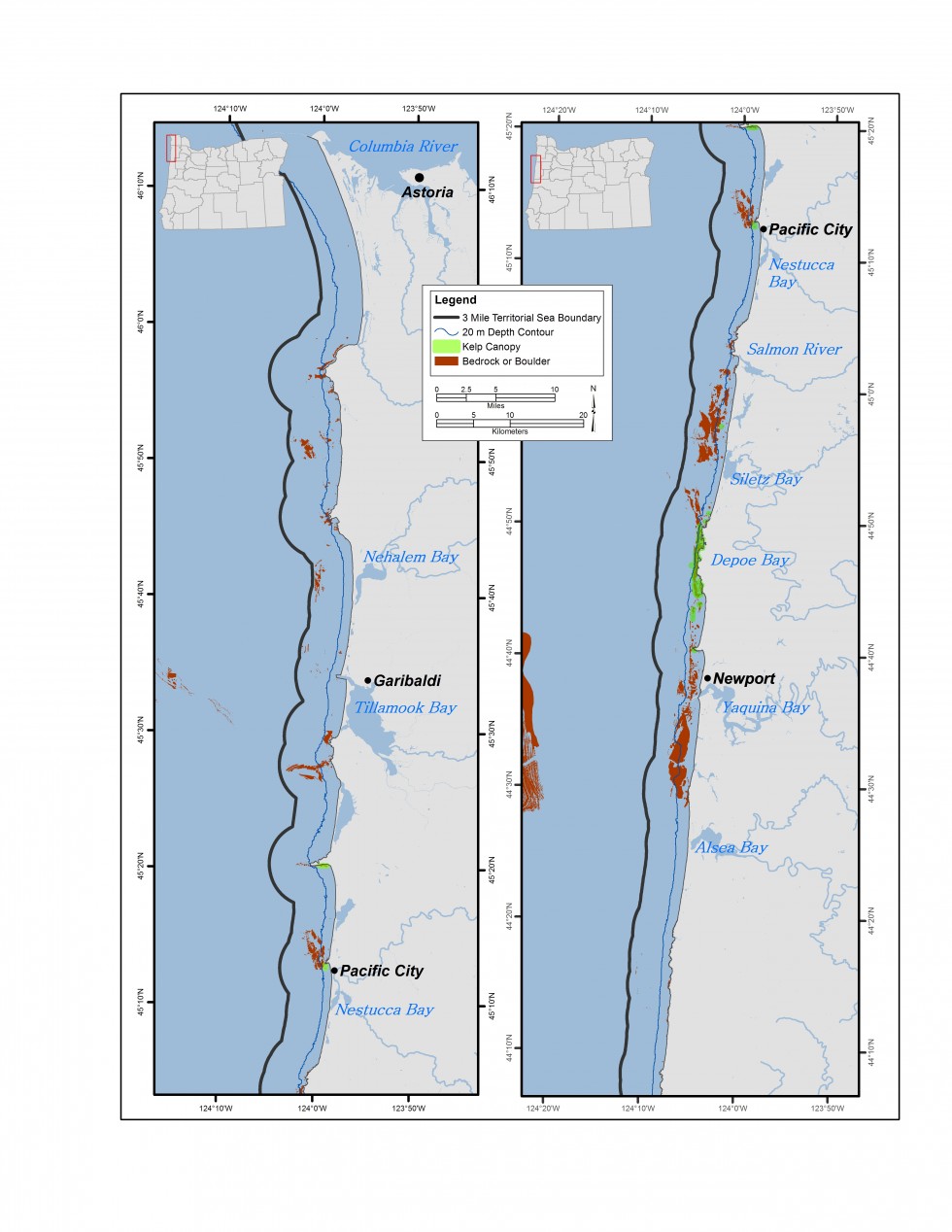
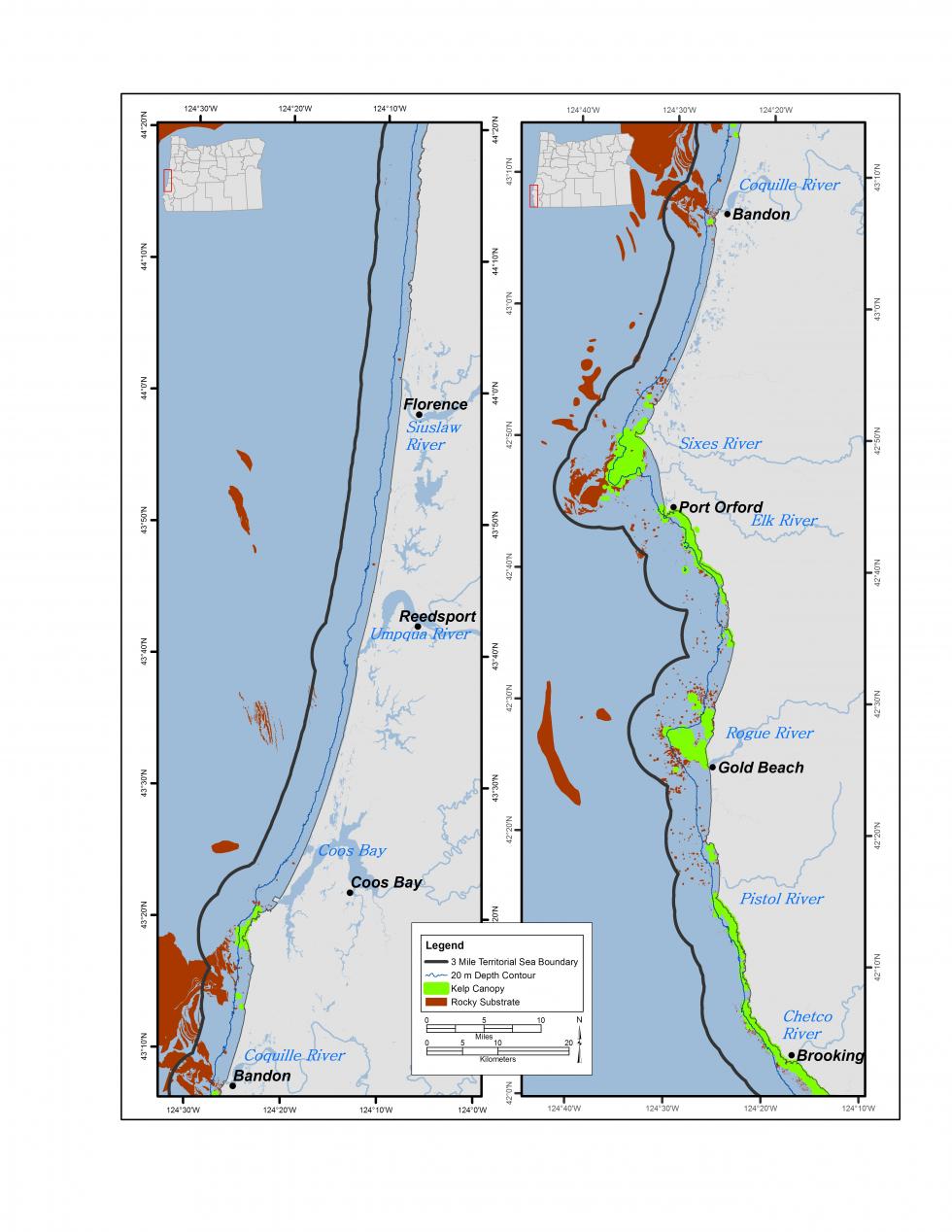
人間の使用
沿岸の岩礁の人間の使用には、漁業、科学研究、観光、その他多くのレクリエーションや産業の追求が含まれます。 多くのタイプのロックフィッシュ種、lingcod、cabezon、およびケルプgreenlingのための商業および娯楽釣はこれまでにこの生息地の第一次人間の使用である。 スキューバダイビングや水中写真は、他のあまり普及している用途の一つです。 商業的な活魚漁業の多くは、浅い沿岸のサンゴ礁で行われます。 レクリエーション釣り人はまた、彼らが利用可能であれば、浅い海岸のサンゴ礁を好む。 沿岸種を対象とした商業漁業の努力は、南海岸では高くなり、北海岸ではレクリエーションの努力がより一般的になる傾向がある。 ユニークな潜在的な商業的使用は、昆布の収穫です。 商業昆布の収穫は、過去に小規模でオレゴン州で数回試みられてきました。 現在、昆布の商業的な収穫はありません。 多くのサンゴ礁は、スキューバダイバー、シーカヤック、ボーター、サーファーによってレクリエーションに使用されています。 広範な昆布ベッドと島を持つサンゴ礁は、沿岸住民や訪問者のための観光やバードウォッチングの機会を提供します。 しかし、多くのサンゴ礁は海面に伸びる特徴を持たないため、多くの人々は水面のすぐ下に存在する豊かな生活に気づいていません。