
Hvordan Replikerer Mitochondia?
Mitokondriell replikering er diagrammed i tegneserien i sidelinjen og vist ovenfor i en elektronmikrograph. Mitokondrier replikerer mye som bakterielle celler. Når de blir for store, gjennomgår de fisjon. Dette innebærer en furrowing av det indre og deretter den ytre membranen som om noen klemte mitokondrionen. Deretter splittet de to datter mitokondriene. Selvfølgelig må mitokondriene først replikere DERES DNA. Dette vil bli diskutert mer detaljert i neste avsnitt. En elektronmikrograf som viser furingsprosessen, er vist i disse figurene. Figuren ovenfor ble tatt Fra Fawcett, En Lærebok I Histologi, Chapman Og Hall, 12. utgave, 1994

noen ganger syntetiseres nye mitokondrier de novo i sentre som er rike på proteiner og polyribosomer som trengs for deres syntese. Elektronmikrografen i figuren ovenfor viser et slikt senter. Det ser ut til at klyngen av mitokondrier sitter i en matrise av proteiner og andre materialer som trengs for produksjonen. Hvordan kan du bevise at materialet i den regionen gjorde mitokondrielle proteiner? Return To Menu
Visse mitokondrielle proteiner er nødvendig før mitokondriene kan dele seg.
Dette har blitt vist i en studie Av Sorgo Og Yaffe, J Cell Bio. 126: 1361-1373, 1994. De viste resultatet av fjerning av et ytre membranprotein fra mitokondrier kalt MDM10. Denne figuren viser resultatene. Mitokondriene er i stand til å ta i komponenter og produsere membraner og matriksenzymer. Fisjon er imidlertid ikke tillatt. Dermed er resultatet en gigantisk mitokondrion. Dette er illstrated i mikrografen nedenfor.

Mitokondrielt DNA og dets funksjon.
Mitokondrier har noe AV SITT EGET DNA, ribosomer, og kan lage mange av sine egne proteiner. DNA er sirkulært og ligger i matrix.in punkterte strukturer kalt «nukleoider». Hver nukleoid kan inneholde 4-5 kopier av mitokondrielt DNA (mrDNA).
Humant mitokondrielt DNA er 16 569 bp; koder for et antall mitokondrielle proteiner
- underenheter 1, 2 og 3 av cytokrom oksidase
- Underenheter 6, 8,9 Av Fo-Atpasen
- Apocytokrom b-underenhet Av CoQH2-Cytokrom c-reduktase
- syv nadh-coq reduktase-underenheter
kjernen koder for de resterende proteinene. Det meste av lipidet importeres (husk forelesningene om lipidtilsetning til membraner). Denne tegneserien fra teksten viser kjernefysisk engasjement. De uthevede etikettene er stoffer som kan brukes til å blokkere prosessen og teste kilden til mitokondrielt protein.

Mitokondrier har også egne ribosomer og tRNA:
- 22 trna
- rna
- 16s
- 12S
- 5S
(Magalhaes, PJ; Andreu, AL, Schon EA, Bevis for tilstedeværelsen av 5 S rRNA i pattedyrs mitokondrier Mol Biol Celle 9: 2375-2382)
Figuren til venstre viser mitokondrielle Ribosomer som granulater i mitokondriene.
teksten sier fortsatt at mitokondrier ikke har 5S rRNA, men den siste studien som er nevnt ovenfor, viser bevis FOR 5S i nøye preparerte mitokondrielle fraksjoner. Disse arbeiderne fant 5S i høyrensede mitokondrier og mitoplaster (mitokondrier uten ytre membran). Konklusjon: 5S rRNA importeres til mitokondrier, men funksjonen er usikker.
Visualisering av mitokondrielt DNA
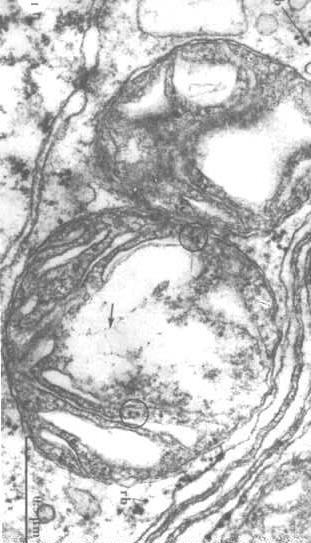
for å visualisere  strukturen av mitokondrielt DNA, må vi trekke ut proteinene i matrisen og avsløre DNA (piler i figuren til høyre).
strukturen av mitokondrielt DNA, må vi trekke ut proteinene i matrisen og avsløre DNA (piler i figuren til høyre).
man kan også se ribosomer i sirklene.
Alternativt kan MAN trekke UT DNA og flyte det på en vannoverflate. Deretter kan den plukkes opp av et plastbelagt rutenett og undersøkes i elektronmikroskopet. Mitokondrielt sirkulært DNA er vist i følgende figur. Denne elektronmikrografen er hentet Fra Fawcett, En Lærebok For Histologi, Chapman Og Hall, 12. utgave, 1994.
Mitokondriell Arv
hos pattedyr er 99,99% av mitokondrielt DNA (mtDNA) arvet fra moren. Dette skyldes at sæden bærer sine mitokondrier rundt en del av halen og har bare ca 100 mitokondrier sammenlignet med 100.000 i oocyten. Etter hvert som cellene utvikler seg, blir flere og flere av mtDNA fra menn fortynnet. Derfor er mindre enn en del i 104 eller 0,01% av mtDNA faderlig. Dette betyr at mutasjoner av mtDNA kan overføres fra mor til barn. Det har også implikasjoner hvis man gjør kloning av pattedyr ved bruk av somatiske celler. Det nukleare DNA ville være fra donorcellen, men mtDNA ville være fra vertscellen. Dette er hvordan dolly sauene ble klonet.
Det er En Gjærstamme, kalt «Petite» som har strukturelt unormale mitokondrier som ikke er i stand til oksidativ fosforylering. Disse mitokondriene har mistet noe ELLER alt DERES DNA. Mitokondriell arv fra gjær er biparental, og begge foreldreceller bidrar til dattercellene når de haploide cellene smelter. Etter meiose og mitose er det tilfeldig fordeling av mitokondrier til datterceller. Hvis fusjonen er med gjær som er petite og gjær som ikke er, vil en viss prosentandel av dattercellene være «petite».
Mutasjoner i pattedyrsmtdna forårsaker sykdommer, fordi det er så kort sekvens og veldig tungt informasjonsinnhold i sekvensen. Den neste foreleseren på mitokondrier i denne serien vil bruke mye tid på mitokondriell genom. Siden hver celle inneholder hundrevis av mitokondrier og tusenvis av kopier av genomet, kan effektene av de muterte mitokondriene fortynnes ut. Som forventet vil de vev eller organer som mest sannsynlig vil bli påvirket, være de som er mest avhengige av oksidativ fosforylering (ATP-produksjon). Hos unge kan det ikke bli plukket opp fordi selv en person med 15% normale mitokondrier kan ha nok til å være sunn. Imidlertid kan aldrende pasienter vise en mer alvorlig sykdom fenotype.
noen eksempler på sykdommer:
- lebers arvelige optiske nevropati( degenerasjon av optisk nerve, ledsaget av økende blindhet): forårsaket av mutasjon til genet som koder for underenhet 4 AV NADH-C0Q reduktase.
- «ragged muskelfibre» assosiert med rykkete bevegelser er forårsaket av mutasjon av mitokondriell lysin tRNA.
- Kaerns-Sayre syndrom: øyefeil, unormal hjerterytme, Degenerasjon Av Sentralnervesystemet. Flere store slettinger i mtDNA.
kan skadet mitokondrielt DNA repareres?
Nåværende studier sier ja.
- Meeusen, S, Tieu, Q, Wong E, Weiss, E, Schieltz, D, Yates, JR, Og Nunnari, J. Mgm101p er en ny komponent av mitochonrial nukleoid som binder DNA og er nødvendig for reparasjon av oksidativt skadet mitokondrielt DNA. J Cell Biol 145: 291-304 (1999)
- Mgm står for «mitokondrie genome maintenance». Det ble oppdaget i gjærceller mens du søkte etter mutanter som forårsaket et temperaturfølsomt tap av mitokondrielt DNA.
- Smeltet Mgm101 til grønt fluorescerende protein og fant at det var lokalisert til punktat» nukleoid » strukturer. Lokalisering overlappet MED DET AV DNA-deteksjonssystemer.
- etter at protein screening fant Mgm101, studerte de hvordan tapet påvirket respiratorisk kompetanse. Klart proteinet var nødvendig for funksjon, men de vet ikke nøyaktig hva dens rolle er på dette punktet.
- Så PÅ COOH terminal-regionen og så at DET var svært grunnleggende. Det antydet At Mgm101p kan ha evnen TIL å binde DNA. Sammenlignet bindingen TIL DNA – cellulosekolonner (ved høye saltforhold) med et ANNET KJENT DNA-bindende protein og bekreftet relativt høy affinitetsbinding av begge proteinene.
Hva skjer med gamle, utslitte mitokondrier?
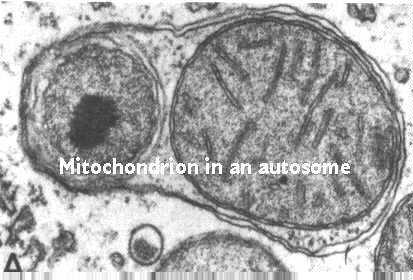
Mitokondrielle tall styres av autofagi. Dette er en prosess hvor lysosomer er involvert i å kontrollere cellekomponenter. Denne Figuren viser prosessen; Den er hentet Fra Fawcett, En Lærebok For Histologi, Chapman And Hall, 12. utgave, 1994.
prosessen begynner ved å pakke endoplasmatiske retikulummembraner rundt mitokondrionen. Deretter kommer vesikler fra Golgi-komplekset og går sammen med autofagisk vakuol. Disse vesiklene inneholder hydrolaser festet til mannose 6 fosfatreseptorer i vesikelmembranene. Lysosomens nettside diskuterer deres funksjon og skjebne. Husk at de smelter sammen med autofagisk vakuol. Syrens pH tillater deretter hydrolasene å bli fjernet fra deres reseptorer. Reseptorene resirkuleres tilbake Til Golgi-komplekset i andre vesikler.
i mellomtiden dannes lysosomet når pH faller og cellene begynner å nedbryte innholdet. Husk at lysosomer er LAMP+, men de bærer IKKE MPR fordi disse har blitt resirkulert Til Golgi-Komplekset. Hva frakk er funnet rundt transport vesikler går til autophagic vacuole?
For mer informasjon, kontakt:
Gwen Childs, Ph. D., FAAA
Professor Og Leder
Institutt For Nevrobiologi Og Utviklingsvitenskap
University Of Arkansas For Medisinske Fag
Little Rock, AR 72205
for spørsmål, kontakt denne e-postadressen: