소개
유아 신경 외측 이영양증은 알려지지 않은 빈도의 상 염색체 열성 희귀 신경 퇴행성 질환입니다. 증상의 발병은 일반적으로 6 개월에서 3 세 사이에 발생합니다. 그 전에 유아는 정상적으로 발달합니다. 이나드의 첫 번째 증상은 정상적인 발달 이정표 또는 발달 이정표에서의 회귀 달성의 둔화 일 수 있습니다(라마나 담 외., 2015). 몸통 근력 저하,사시 및 안진 증은 질병의 초기 증상입니다(그레고리 외., 2017). 이 질병의 진행은 빠르며 진행됨에 따라 더 많은 획득 기술이 손실됩니다. 근육은 곧 저장성이되어 나중에 경련이됩니다(리바이스와 피나치,2014). 결국 모든 자발적인 근육 조절이 상실됩니다. 근육 약화는 또한 수유 및 호흡에 어려움을 초래할 수 있습니다. 안진 증 외에도 일부 어린이는 시력 상실을 경험합니다. 인지 기능이 점차 상실되고 치매가 발생합니다. 수명은 일반적으로 5~10 년(그레고리 외. 1993;자르 딤 외. 2004;맥컬리와 샌즈,2009).274>
분자 병리학
영아 신경 외측 이영양증은 비정형 후기 발병 신경 외측 이영양증(아나 드)및 근긴장 이상 파킨슨증 복합체를 포함하는 신경 퇴행성 장애의 계열에 속한다. 대부분의 경우 이나드는 단백질 생성물의 촉매 활성에 영향을 미치는 동형 접합체 또는 화합물 이형 접합체 돌연변이와 관련이 있습니다(엥겔 외., 2010). 상기 유전자는 리파아제 및 7 개의 안키린 반복 함유 도메인(탕 외)과 함께 칼슘-독립 포스포리파아제 2 단백질(탕 외)을 통해 그룹을 코딩한다., 1997). 또한,이 물질은 인지질,활성 지방산 및 리소 인지질을 생성 할 수 있습니다.
미토콘드리아 내막 내의 인지질은 불포화 지방산,특히 카디오리핀이 풍부하다., 2006). 이 불포화 지방산은 미토콘드리아(머피,2009)에 의해 생성 된 풍부한 반응성 산소 종에 특히 취약하여 미토콘드리아의 내부 막에 과산화 된 인지질을 생성합니다. 미토콘드리아(윌리엄스와 고트립,2002;리오우 외. 2005)리모델링 된 인지질(발사 신드 외.2005)에 이르는 인지질의 위치에있는 과산화 지방산의 가수 분해에 대한 수요 증가와 일치.,1995;자오 등., 2010). 미토콘드리아 내부 막 무결성이 손상됩니다. 또한 축삭에 국소화된다. 2005;셀레즈네프 외. 2006)인지질 리모델링에 대한 현지화 된 수요가 증가했음을 나타냅니다. 두뇌에 있는 그런 축적의 표현은 표절 26 를 포함하는 동일한 근본적인 분자 병인을 위한 각종 이름 귀착된 기초 신경절과 같은 중요한 두뇌 지역에 유일합니다(멘나즈,2016 년;나시프 외., 2016).
녹아웃 마우스의 뉴런의 미세구조 분석은 이 분자 병리와 일치한다. 분기 및 관상 크리스타를 갖는 미토콘드리아,퇴화 된 크리스타를 갖는 미토콘드리아,세포 골격 붕괴를 갖는 축삭 및 축삭 말단에서 부분 막 손실이 관찰되었다(벡 외., 2011). 현미경 수준에서 이러한 기능은 중추 또는 말초 신경계의 시냅스 전 말단(그림 1)에서 축삭 부종 및 회전 타원체로 나타납니다.
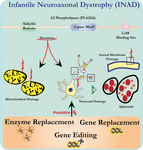
그림 1. 이 질환은 신경 퇴행성 장애,신경 퇴행성 장애,신경 퇴행성 장애,신경 퇴행성 장애,신경 퇴행성 장애,신경 퇴행성 장애,신경 퇴행성 장애,신경 퇴행성 장애,신경 퇴행성 장애,신경 퇴행성 장애,신경 퇴행성 장애,신경 퇴행성 장애. 미토콘드리아 및 축삭 막 결손을 유발하는 기능 장애를 유발합니다. 이러한 결함은 축삭 팽창 및 시냅스 전 회전 타원체의 축적으로 시각화 된 신경 손상을 유발합니다. 기능을 복원하기 위한 효소 대체,유전자 대체 또는 결함이 있는 판 2 를 교정하기 위한 편집이 제안된 치료 전략이다.
분자 진단 및 희귀 질환 환자 역량 강화
다음 세대 시퀀싱의 가용성 이전에 특정 임상,전기 생리 학적,및 영상 기능 외에도 피부 생 검 축 삭 부종과 중앙 또는 말 초 신 경계에 사전 시 냅 스 터미널에서 구상 체를 보여주는 했다(그레고리 등의 확인에 대 한 진단 기준. 2017;요오디스외., 2017). 종종 진단을 확인하기 위해 여러 번의 생검이 필요했습니다. 가족은 일반적으로 진단을 위해 수년을 기다렸다. 유전자 및 게놈 시퀀싱의 감소 비용,진단 되지 않은 신경 질환에 대 한 진단 실험실과 테스트 대상된 유전자 패널의 가용성 및 유전자 진단의 가용성의 의사의 증가 인식 가족 받고 진단 더 급속 하 게;때로는 첫 번째 증상이 나타나는 1 년 이내. 이 가족의 아이들은 아직 어리고 가족들은 과학자들과 협력하여 자녀의 질병 치료법을 찾도록 동기를 부여합니다. 연구 과정에 자금을 지원하기 위해,이나드 환자의 부모 그룹이 인더큐어재단을 구성했습니다. 재단은 연구를위한 상당한 자금을 제기하고 연구 보조금의 수여에서 그들을 안내하는 희귀 유전체학 연구소와 제휴한다. 2016 년 10/22 가족(45.45%)이 유전자 코딩 영역에서 돌연변이가 부족하다는 것을 발견했습니다., 2016). 유전자 변이를 식별 하기 위해 전체 유전자 시퀀싱 필요 미래의 분자 진단 노력을 강조의 코딩 영역에서 해로운 돌연변이를 식별 하는 실패 또한,플라 2 제 6 과 관련된 돌연변이가 관찰되지 않은 경우,그 질병의 원인은 플라 2 제 6 과 관련된 돌연변이가 아닌 다른 유전자의 돌연변이에 기인할 수 있음을 시사하며,이는 탐구되어야 한다.
가능한 치료법
결함있는 효소를 포함하는 이나드와 같은 희귀 질환에 대해 고려되는 대부분의 치료법은 효소 대체,유전자 대체 또는 유전자 교정이다. 효소 결핍이 열성 유전 적 결함에 의해 야기 될 때,효소 대체 또는 보충이 문제를 해결할 수 있다고 가정된다(스미스 외. 2012;유와이만,2016). 그러나,정확한 유전자 또는 효소를 제공하는 치료가 나드 표현형을 구출할 것이라는 것을 증명하는 실험은 아직 실행되고 시험될 것입니다.뇌가 이나드에 영향을 받는 1 차 기관이기 때문에,이나드에 대한 효소 대체 요법은 대부분 효소를 뇌에 주입해야 할 것이다. 2017 년 바이오마린은 트리펩티딜 펩티다아제 1(세리포나아제 알파)을 배튼 질환의 근본 원인,트리펩티딜 펩티다아제의 결핍에 대한 치료제로서 리소좀 효소(미국 식품의약품안전청,2017)의 승인을 받았다. 트리 펩티딜 펩 티다 제 1(세리 포나 제 알파; 뇌척수액(뇌척수액)에 직접 투여되는 첫 번째 뇌척수액(뇌척수액)입니다. 뇌척수액으로 투여될 수 있는 다른 뇌척수액 약물은 임상시험에 있다(자르 딤 외. 2004;맥컬리와 샌즈,2009).
배튼 병 환자의 보행 능력에 대한 트리 펩티딜 펩티다제 1(세리포나제 알파)의 효능은 뇌에 영향을 미치는 질병에 대한 효소 대체 요법이 이론적으로 가능하다는 것을 입증한다. 이나드에 대한 미토콘드리아로 대체 효소를 표적화하는 것은 고셔병의 경우처럼 오랫동안 4 에르트에 의해 이미 성공적이었던 리소좀 표적화보다 더 어려울 것이다. 따라서,아직 해결해야 할 많은 생물학적 문제들이 있다.
•미토콘드리아에 플라투그 6 을 국소화하기 위해 어떤 메커니즘을 사용할 수 있는가?
•축삭에 국소화 할 필요가 있습니까?
•말초신경계를 표적으로 하는 특별한 필요가 있는가?
•일부 단백질 결핍을 구할 수 있습니까,아니면 전체 단백질이 필요합니까?
•뇌척수액으로 유전자 생성물을 전달하기 위한 적절한 용매는 무엇인가?
•만약 100%순수하지 않다면,그 오염물질은 무엇이며 안전한가?
•중추 신경계에 효소 주입으로 인한 일반화 된 혈관 내 합병증이 있습니까? 주입 용량이 그러한 합병증에 어떤 관련이 있습니까?
추가 문제는 효소 대체 요법이 심실 내 카테터의 초기 배치와 빈번한 주입을 필요로한다는 것입니다. 카테터의 배치는 마취가 필요하며 주입은 환자의 협력에 따라 마취가 필요할 수 있습니다. 소아 신경 학자와 적절 한 정보를 가진 아 드 환자에 마 취를 사용 하 여 관련 아 드와 아 드와 아 드의 부모의 문제를 해결 추가 노력이 필요 합니다.유전자 치료/유전자 대체
인간 플라 투지 유전자는 약 70 킬로바이트이며,17 개의 엑손과 2 개의 대체 엑손이 있다. 가장 긴 단백질 코딩 성적 증명서는,그러나,단지 3.3 킬로바이트이며,단백질 코딩 서열은 쉽게 바이러스 벡터화물로 포장 할 수 있습니다 그냥 이상 2.4 킬로바이트입니다. 우리는 먼저 중추 신경계에서 전임상 데이터를 가진 다른 리소좀 저장 장애를 탐구 한 다음 인애드와 비교할 것입니다. 전임상 모델 유전자 치료 연구를 겪고 있는 중추 신경계 질환의 다른 전임상 모델을 관찰 하는 중요 한 이유는 분야에서 상당한 지식 격차입니다. 이전에,마우스의 전임상 연구는 바이러스 벡터 안에 설파미다제 유전자의 올바른 버전을 포장하여 리소좀 저장 장애 점액 다당류 유형 이아(의원-이아)에 대한 유전자 치료를 해결(소렌티노 외., 2013). 이 연구는 혈액 뇌 장벽에 대한 이해의 증가를 이용했습니다.이 장벽은 혈액에서 중추 신경계로의 큰 분자의 수송을 능동적으로 조절하는 과정을 통해 트랜스 사이토 시스(파드리지,2005 비,소렌 티노 및 프랄디,2016).
트랜스 사이토 시스는 리간드 특이 적 수용체(예:인슐린 수용체,트랜스페린 수용체 및 저밀도 지단백질 수용체 등)에 의해 매개되는 내강 측에서 리간드의 엔도 사이토 시스를 포함한다.모세 혈관 내피에 농축 됨(파 드리지,2005 에이). 이것은 내피 세포질을 통해 세포 내화 된화물의 이동과 마지막으로 뇌(뇌)쪽에서 세포 외 세포 증이 뒤따라서화물을 효과적으로 전달합니다(파드리지,2002). 2013 년 11 월 1 일,서울시 강남구 테헤란로 15 길 11(우)1123-1111 사업자등록번호:120-81-00000 통신판매업신고번호:제 2013-서울강남-0001 호 대표전화:02-3100-0000 통신판매업신고번호:제 2013-서울강남-0001 호 대표전화:02-3100-0000 통신판매업신고번호:제 2013-서울강남-0001 호 대표전화:02-3100-0000, 2013). 키메라 설파 미다 제를 코딩하는 바이러스 벡터화물은 간을 표적으로하는 아데노 관련 바이러스(에이브)혈청 형 2/8 에로드되었습니다(소렌 티노 및 프랄디,2016). 따라서 간은 키메라-설파 미다 제의 지속적인 공급을 제공하는 내부 공장으로 작용하여 간 유전자 치료 후 7 개월 후에도 뇌 설파 미다 제 활성이 10-15%증가했습니다. 뇌 설파 미다 제 활성 수준의 증가는 마우스 모델의 뇌 병리학 및 행동 결과를 정량화 할 수있는 개선으로 이어졌다., 2013).
이나드에 대해 유사한 연구가 수행된다면,이나드에 대한 효소 대체에 관한 몇 가지 중요한 질문에 답할 것이다. 또한,혈관내 또는 뇌척수액 내 투여는 중추 신경계를 직접 표적으로 할 수 있다.,2017;로카 외., 2017). 그러나,이나드 환자들이 그들의 표절 26 유전자에 가지고 있는 여러 돌연변이는 유전자 치료에 독특한 도전을 제공한다. 그러나 2-3 킬로바이트의 플라 2 유전자 유전자의 크기 바이러스 벡터에 삽입에 대 한 문제를 제기 하지 해야,,플라 2 유전자 수정에서 규제 합병증 예측하기 어렵다. 규제 합병증은 특히 유전자 치료가 표적 조직 내에서 적절하게 국소화 될 수없는 경우 통제 할 수 없게됩니다. 일부 돌연변이 제품이 야생 형 표절에 지배적 인 부정적인 것으로 판명되면 효소 대체 요법도 문제가 될 수 있습니다 26.
이나드에 대한 가능한 치료법의 대부분의 과제는 그것이 매우 드문 질병이라는 사실에서 비롯된다. 그러나,이나드가 알려진 유전적 병인을 가지고 있다는 사실은 가능한 치료법에 대한 길을 제공한다. 또한,이나드에 대한 성공적인 치료는 고아 약물 상태 및이나드가 드문 질환이기 때문에 그것이 얻는 모든 보호를 받게됩니다. 따라서 앞서 언급 한 모든 도전에도 불구하고 고아 약물 상태는 유전 적 원인이 이미 알려진 경우는 말할 것도없고 희귀 질환 연구자 및 생명 공학 산업에 대한 강력한 인센티브를 제공합니다.유전자/염기 편집
2017 년 현재,적어도 277 개의 미스 센스 돌연변이가 인간 유전자에서 관찰되었다., 2016). 프레임 이동,인델,말도 안되는 돌연변이 및 스플 라이스 부위의 돌연변이가 포함됩니다(모건 외., 2006). 따라서 표적 세포 집단에 있는 유전자를 정정하거나 돌연변이된 유전자 기초를 정정하는 것은 매력적인 치료 도로입니다. 수년에 걸쳐,유전자 편집을위한 몇 가지 도구가 개발되었으며,크리스퍼/카스 9 기반 게놈 편집은 2018 에서 계획된 임상 시험과 함께 획기적인 기술로 환영 받고 있습니다. 이러한 기술은 새로운 유전자 서열의 삽입 또는 해로운 유전자 염기의 교정을 위한 공간을 만들기 위해 지정된 방식으로 스트랜드를 절단할 수 있는 유전자 결합 단백질을 활용한다., 2015). 이미 여러 전임상 질환 모델(예:뒤신 근이영양증,간 대사성 질환 등)에서 놀라운 치료 결과를 얻었습니다.,)(다이 외., 2016). 또한,인간의 변이에 대한 이해는 유전자 치료에 대한 새로운 도전을 제기하고 유전자 편집의 진정한 개인화를 향한 혁신을 주도하고 있습니다(레저드 외. 2017;스콧과 장,2017). 신경 세포 및 기타 신경계 세포의 결함 유전자를 대체하기 위한 새로운 기술(니시야마 외)은 유전자 편집 기술을 새로운 국경으로 추진하고 있습니다., 2017).
결론
유아 신경성 이영양증은 특정 이환율과 사망률을 가진 심각한 신경 퇴행성 질환입니다. 이 희귀 질환은 차세대 요법의 사용 가능한 모드를 재 검증하고 새로운 치료법을 생성 할 수있는 흥미로운 기회를 제공합니다. 인애드의 임상 진단 기준에 대한 새로운 합동은 향상된 분자 진단을 향한 자극을 제공 할 것으로 예상됩니다. 이 진전은 우리로 하여금 갑상선 기능 항진증 환자의 삶의 질을 정량화 할 수있는 개선을 제공하고 질병 진행을 지연 시키거나 개선 할 수있는 저렴한 치료법을 개발하게 할 것으로 예상됩니다. 우리는 배튼 병과 뮤코 다당류의 성공 사례를 강조하여 이나 드 요법을 현실로 만들기 위해 올바른 질문을 할 수있는 영감을 제공했습니다. 또한,크리스퍼/카스 9 기반 유전자 편집에 대한 바이러스 및 비 바이러스 접근법의 성장은 또한 인애드에 대한 새로운 치료 방법을 열어야한다.
저자 기여
인민위원회는 예비 초안을 준비. 시니어와 교류는 편집 및 원고에 더 많은 섹션을 추가. 원고를 편집했습니다.
이해 상충 성명
저자는 연구가 잠재적 인 이해 상충으로 해석 될 수있는 상업적 또는 재정적 관계가없는 상태에서 수행되었다고 선언합니다.2010 년 11 월 15 일,2010 년 12 월 15 일,2010 년 12 월 15 일,2010 년 12 월 15 일,2010 년 12 월 15 일,2010 년 12 월 15 일,2010 년 12 월 15 일,2010 년 12 월 15 일,2010 년 12 월 15 일,2010 년 12 월 15 일,2010 년 12 월 15 일,2010 년 칼슘 독립적 인 포스 포 리파아제의 억제는 아라키돈 산 혼입 및 인지질 리모델링을 방지합니다. 프록 내틀 아카드 과학. 미국 92,8527-8531. 도이:10.1073/나스.92.18.8527
PubMed Abstract | CrossRef Full Text | Google Scholar
Beck, G., Sugiura, Y., Shinzawa, K., Kato, S., Setou, M., Tsujimoto, Y., et al. (2011). Neuroaxonal dystrophy in calcium-independent phospholipase A2beta deficiency results from insufficient remodeling and degeneration of mitochondrial and presynaptic membranes. J. Neurosci. 31, 11411–11420. doi: 10.1523/JNEUROSCI.0345-11.2011
PubMed Abstract | CrossRef Full Text | Google Scholar
Bey, K., Ciron, C., Dubreil, L., Deniaud, J., Ledevin, M., Cristini, J., et al. (2017). 신경 질환의 유전자 치료를 위해 단일 좌초 된 단일 좌초 정맥 내 주입에 의해 성인 쥐에서 효율적인 중추 신경계 표적화. 진 거기. 24:325. 도이:10.1038/있다.2017.18
PubMed 추상|CrossRef 전체 텍스트|Google 학술검색
Dai,W.-J.,주홍,L.Y.,Yan,Z-Y.,쑤,Y.,Wang,Q.L.,Lu,X-J.,et al. (2016). 생체 내 유전자 치료를위한 크리스퍼-카스 9:약속과 장애물. 몰 거기 핵산 5:전자 349. 도이:10.1038/미트 나.2016.58
출판 초록|크로스레프 전체 텍스트|구글 학자
엥겔,엘에이,징,징,오브라이언,디이 2010 년 11 월 15 일,2010 년 11 월 15 일. 유아 신경성 이영양증과 관련된 돌연변이에 의해 손상되지만 근긴장 이상-파킨슨증은 아닙니다. 2015 년 11 월 15 일. 도이:10.1371/저널.폰0012897
펍드 초록/교차 참조 전체 텍스트
2015 년 12 월 1 일,2015 년 12 월 1 일,2015 년 12 월 1 일,2015 년 12 월 1 일,2015 년 12 월 1 일,2015 년 12 월 1 일,2015 년 12 월 1 일,2015 년 12 월 1 일. 2015 년 11 월 1 일,서울시 강남구 테헤란로 101(우)1001 호(우)1001 호(우)1001 호(우)1001 호(우)1001 호(우)1001 호(우)1001 호(우)1001 호
구글 학자
2018 년 11 월 1 일-2018 년 12 월 1 일-2018 년 12 월 1 일-2018 년 12 월 1 일-2018 년 12 월 1 일, and Hayflick, S. J. (2017). PLA2G6-Associated Neurodegeneration. Available at: https://www.ncbi.nlm.nih.gov/books/NBK1675/
Google Scholar
Iodice, A., Spagnoli, C., Salerno, G. G., Frattini, D., Bertani, G., Bergonzini, P., et al. (2017). Infantile neuroaxonal dystrophy and PLA2G6-associated neurodegeneration: an update for the diagnosis. Brain Dev. 39, 93–100. doi: 10.1016/j.braindev.2016.08.012
PubMed Abstract | CrossRef Full Text | Google Scholar
Jardim, L., Vedolin, L., Schwartz, I. V., Burin, M. G., Cecchin, C., Kalakun, L., et al. (2004). 파브리 병에 중추 신경계 관련:효소 대체 요법 12 개월 전후의 임상 및 영상 연구. 제이.상속. 메타. 아뇨 27, 229–240. 도이:10.1023/비:볼리.0000028794.04349.91
PubMed 추상|CrossRef 전체 텍스트|Google 학술검색
Kapoor,S.,Shah,M.H.,싱,N.,오히려,M.I.,Bhat,V.,난,S.,et al. (2016). 유아 신경 외음부 이영양증,비정형 후기 발병 신경 외음부 이영양증 및 근긴장 이상 파킨슨증 복합체가있는 22 명의 인도 가정에서 플라 2 지 6 의 유전 분석. 11:155605. 도이:10.1371/저널.폰0155605
출판 초록|크로스 레프 전체 텍스트|구글 학자
2015 년 11 월 15 일,2015 년 11 월 15 일,2015 년 11 월 15 일,2015 년 11 월 15 일,2015 년 11 월 15 일. 유전자 편집 기술의 전달 및 치료 응용. 2015 년 11 월 15 일-2015 년 11 월 15 일 지능. 제약회사 494, 180–194. 도이:10.1016/제이.2015.08.029
PubMed 추상|CrossRef 전체 텍스트|Google 학술검색
Lek,M.,Karczewski,K.,Minikel,E.,Samocha,K.E.,은행,E.,Fennell,T.,et al. (2016). 60,706 인간의 단백질 코딩 유전 적 변이 분석. 자연 536,285-291. 도이:10.1038/nature19057
PubMed 추상|CrossRef 전체 텍스트|Google 학술검색
Lessard,S.,Francioli,L.,Alfoldi,J.,tardif 의,J.C.,Ellinor,P.T.,맥아더,D.G.,et al. (2017). 인간 유전 변이는 치료적으로 연루된 유전자좌에서 크리스퍼-카스 9 온-및 오프-타겟팅 특이성을 변경한다. 프록 내틀 아카드 과학. 미국 14,전자 11257-전자 11266. 도이:10.1073/나스.1714640114
출판 초록/크로스 레프 전체 텍스트|구글 학자
2014. 뇌 철분 축적을 통한 신경 퇴화: 병원성 메커니즘에 대한 업데이트. 전면. 파마솔. 5:99. 도이:10.3389/파르.2014.00099
크로스레프 전체 텍스트/구글 학자
본 발명의 다른 실시예는,본 발명의 다른 실시예에 따른 실시예에 따른 실시예에 따른 실시예에 따른 실시예에 따른 실시예에 따른 실시예에 따른 실시예에 따른 실시예에 관한 것이다. (2005). 인간 암세포에서 시클로 옥 시게나 제 -2 및 칼슘 독립적 인 포스 포 리파아제의 미토콘드리아 국소화:세포 사멸 저항의 함축. 특급. 세포 레.306,75-84. 10.1016/제이.2005.01.011
출판 초록|크로스레프 전체 텍스트|구글 학자
2009 년 12 월 15 일.2009 년 12 월 15 일. 리소좀 저장 질환에 대한 유망한 중추 신경계 지시 효소 대체 요법. 특급. 뉴롤이야 218, 5–8. 도이:10.1016/제이.2009.03.040
출판 초록|크로스레프 전체 텍스트|구글 학자
(2016). 뇌 철 축적과 함께 신경 퇴행(엔 비아)이전에 할레 보덴-스패 츠 질병. 2018 년 물리학입니다 인도 64:132.
구글 학자
이 경우,상기 제 1 항은 제 2 항 및 제 2 항과 제 2 항 및 제 2 항과 제 2 항 및 제 2 항과 제 2 항 및 제 2 항과 제 2 항 및 제 2 항과 제 2 항 및 제 2 항과 제 2 항 및 제 2 항과 제 2 항 및 제 2 항과 제 2 항 및 제 2 항과 제 2 항 및 제 2 항과 제 2 항 및 제 2 항 (2006). 뇌 철분이 높은 신경 퇴행성 질환에서 돌연변이가 발생합니다. 냇 제네트 38, 752–754. 2018 년 10 월 10 일(토)~2018 년 10 월 10 일(일)~2018 년 10 월 10 일(일)~2018 년 10 월 10 일(일)~2018 년 10 월 10 일(일)
2009. 미토콘드리아가 반응성 산소 종을 생성하는 방법. 바이오켐 417,1-13. doi:10.1042/BJ20081386
PubMed 추상|CrossRef 전체 텍스트|Google 학술검색
Nassif,D.,페레이라,J.S.,스,M.,카피통 나,C.,그리고 파리 아,A.(2016). 뇌 철분 축적을 통한 신경 퇴화:사례 보고서. 데멘트요 신경 정신병. 10, 160–164. doi:10.1590/S1980-5764-2016DN1002014
PubMed 추상|CrossRef 전체 텍스트|Google 학술검색
니시야마,J.,삼국,T.,야스다,R.(2017 년). 포유류 뇌의 유사 분열 및 분열 후 세포에서 상 동성 지향 수리를 통한 바이러스 매개 게놈 편집. 뉴런 96,7555–768 이요5. 도이:10.1016/제이.2017.10.004
출판 초록|크로스레프 전체 텍스트|구글 학자
2005 년 11 월 15 일,2005 년 12 월 15 일,2005 년 12 월 15 일,2005 년 12 월 15 일,2005 년 12 월 15 일,2005 년 12 월 15 일. 원숭이 뇌에서 칼슘 독립 포스 포 리파제 2 의 분포. 제이 뉴로시톨 34, 447–458. 도이:10.1007/11068-006-8730-4
째챈쨩처쨀짼쨉쨉짹쨀째철쩔첩쨩
2002. 분자 트로이 목마를 가진 두뇌에 표적으로 하는 약 그리고 유전자. 냇 마약 디스코프 목사. 1, 131–139. 2018 년 10 월 10 일(토)~2018 년 10 월 10 일(일)~2018 년 10 월 10 일(일)~2018 년 10 월 10 일(일)~2018 년 10 월 10 일(일)
2010 년 11 월 15 일. 혈액 뇌 장벽의 분자 생물학. 몰 바이오 테크놀입니다. 30, 57–70. 도이:10.1385/메가바이트:30:1:057
크로스 레프 전체 텍스트/구글 학자
2005 년 11 월 15 일. 혈액-뇌 장벽:뇌 약물 개발의 병목 현상. 뉴 록스 2,3-14.
구글 학자
이 경우,본 발명의 실시예에 따라,본 발명의 실시예에 따른 실시예에 따른 실시예에 따른 실시예가 결정된다. (2015). 칼슘 독립적 인 포스 포 리파제는 생물학적 과정과 질병에서 그 역할을합니다. 56,1643-1668. 10.1194R058701
PubMed 추상|CrossRef 전체 텍스트|Google 학술검색
Roca,C.,Motas,S.,Marcó,S.,Ribera,A.,Sánchez,V.,Sánchez,X.,et al. (2017). 무코 다당류 증 유형의 새로운 마우스 모델에서 아브 매개 유전자 치료에 의한 질병 교정. 흠. 몰 제네트 26, 1535–1551. doi:10.1093/hmg/ddx058
PubMed 추상|CrossRef 전체 텍스트|Google 학술검색
Scott,D.A.,및 장,에프(2017 년). 크리스퍼 기반 치료 게놈 편집에서 인간 유전 적 변이의 시사점. 냇 메드. 23:1095. 도이:10.1038/나노.4377
펍메드 초록|크로스레프 전체 텍스트|구글 학자
2018 년 11 월 1 일 2006. 칼슘 독립적 인 포스 포 리파제 2 는 스타 우로 스포린에 의한 세포 사멸 유도 동안 미토콘드리아를 국소화하고 보호합니다. 제이비올 켐 281, 22275–22288. 도이:10.1074/제이비씨.2018 년 11 월 1 일,2018 년 11 월 1 일,2018 년 11 월 1 일
2012 년 11 월 15 일,2013 년 11 월 15 일,2013 년 11 월 15 일,2013 년 11 월 15 일,2013 년 11 월 15 일. 열성 형태의 유전 된 망막 영양 장애에 대한 유전자 보충 요법. 진 거기. 19, 154–161. 도이:10.1038/있다.2011.161
출판 초록|크로스레프 전체 텍스트/구글 학자
소렌 티노,엔. C.,D Orsi,L.,Sambri,I.,노스코,E.,모나코,C.,Spampanato,C.,et al. (2013). 혈액-뇌 장벽을 통과하도록 설계된 고도로 분비 된 설파 미다 제는 뮤코 다당류 유형 아이 아를 가진 마우스의 뇌 병변을 교정합니다. 엠보 몰 메드. 5, 675–690. 도이:10.1002/음.201202083
출판 초록/크로스레프 전체 텍스트/구글 학자
2016 년 11 월 15 일.2016 년 11 월 15 일. 의원의 뇌 타겟팅. 소아과 내분비. 요한계시록 13 장 1), 630–638.
구글 학자
2018 년 11 월 1 일-2018 년 11 월 1 일-2018 년 11 월 1 일-2018 년 11 월 1 일-2018 년 11 월 1 일 2010 년 11 월 1 일. 새로운 세포질 칼슘 독립 포스 포 리파제 대답 2 에는 8 개의 안 키린 모티브가 포함되어 있습니다. 제이비올 켐 272, 8567–8575. 도이:10.1074/제이비씨.272.13.8567
출판 초록|크로스 레프 전체 텍스트|구글 학자
미국 식품 의약품 안전청(2017). 식약청은 고정편 질환의 형태에 대한 첫 번째 치료를 승인합니다. 에서 사용 가능: https://www.fda.gov/NewsEvents/Newsroom/PressAnnouncements/ucm555613.htm
2010 년 11 월 15 일,2010 년 11 월 15 일. 미토콘드리아 인지질 손실을 약화시키고 심장 보호 작용을합니다. 바이오켐 제 362 조(면 1),23-32. doi:10.1042/bj3620023
PubMed 추상|CrossRef 전체 텍스트|Google 학술검색
Yu-와이-남자,P.(2016). 유전 된 신경 퇴행성 질환에 대한 유전자 조작:신화 또는 현실? 브롬 제 1 조(목적) 100:1322. 도이:10.1136/비요프안솔-2015-308329
째챈쨩처쨀짼쨉쨉짹쨀째철쩔첩쨩
자오,지,장,엑스,자오,씨,최,제이,시,제이,송,케이,외. (2010). 포스 포 리파아제를 통한 그룹 별 췌장 베타 세포 보호 미토콘드리아 막 과산화의(2)매개 수리. 내분비학 151,3038-3048. 도이:10.1210/엔.2010-0016
퍼블릭 초록|크로스레프 전체 텍스트|구글 학자