소개
뉴로 트로 핀 신경 성장 인자는 표적 신경 분포,세포 분화 및 신경 생존(대형 등)을 포함하여 발달하고 성숙한 포유류 신경계에서 몇 가지 중요한 과정에 관여합니다.,1986;리 외. 1995;소프로뉴 외.,2001;래드 외., 2003). 성인 뇌에서,핵발전소는 기초 전뇌 콜린성 시스템의 유지 및 기능에 핵심적인 역할을(젊은이 외., 2003). 기저 전뇌 콜린성 돌기,즉 해마와 피질(코르 싱 외. 이 경우,피질 내 피질 수용체는 기저 전뇌 피질 투영(코다워 외)에 주로 국한된다. 1988;소프로뉴 외. 2001;로시 외., 2002). 신경 성장 인자 신호는 성인기 동안 콜린성 피질 투영을 유지합니다(첸 외. 1997;데베이르 외. 이러한 뉴런의 퇴행은 축삭의 절단에 따른 이들 뉴런의 퇴행을 방지 할 수있다(헤프티,1986;투신 스키 외. 1990;코르다워 외., 1994). 또한,아세틸콜린은 기초 전뇌 배양에서 아세틸콜린의 방출을 향상시킵니다. 콜린성 유전자 궤적의 표현형 제어를 통해 콜린성 기능을 조절할 수 있습니다. 1993;후진타오 외., 1997).
기저부 전뇌 콜린성 시스템은 피질에서 학습 및 경험 의존적 가소성에 영향을 미친다(바킨 및 와인 버거,1996;킬가드 및 메르 체니 치,1998;코너 외. 2003 년,2005 년),그리고 기초 전뇌 매개 학습 및 가소성을 지원하고 향상시킬 수 있다는 가설이 있습니다. 예를 들어,편협한 피질에서 편협한 피질에서 편협한 피질 항체의 반복 주사를 통해 편협한 피질 신호 전달의 봉쇄는 국소 콜린성 신경 분포를 감소시키고 콜린성 입력의 완전성,조절 된 맛 혐오 및 억제 회피 학습에 의존하는 두 가지 과정의 획득을 방해합니다(구티에레즈 외., 1997). 코너 등의 최근 연구. (2009)공간 학습 및 해 마 장기 강화(중격 핵,해 마에 콜린 성 신경 분포의 기본 소스로의 주입 다음 향상 된 시연.
의 초점 응용 프로그램 또한 쥐의 배럴 피 질에서 로컬 수염 표현의 급속 한 확장을 유도할 수 있다. 이 과정은 기저부 전뇌의 콜린성 투영에 의존합니다. 콜린성 의존성 기능 재구성을 자극 할 수 있음을 시사한다. 따라서,기초 전뇌 콜린성 세포에 대한 장기적인 영양 효과 외에도,단기간 회로 리모델링에 참여할 수 있습니다. 이”이중 행동”가설은 구성 및 활동 의존적 방출 메커니즘의 존재에 의해 뒷받침된다. 2003),잠재적으로 장기 영양 및 단기 조절 효과의 기초. 또한,역행(장기)및 로컬(단기)신호 경로(황 및 라이하르트,2003)에 대 한 관찰 되었습니다.
세포 표현 NGF 에서 발견되었습 cortex,소뇌,해마,시상가,기저 개 및 뇌간(담고 Isackson,1989;Maisonpierre et al. 1990;아이삭슨 외. 1991;코너 및 바론,1992;하야시 외. 1993;머프슨 외.,1994;장 외., 2007). 성상세포와 다른 신경교세포에서 관찰된 것으로 알려져 있지만,대부분의 피질 신경교세포는 뉴런에 의해 생성된다., 2001). 그러나 콜린성 피질 투영은 피질에서 흥분성 및 억제성 뉴런을 모두 자극하는 것으로 나타났습니다., 1999). 해마,선조체 및 기저 전뇌와 같은 외 피질 영역에서,핵분열 생성은 주로 가바 에르 억제 뉴런에 국한됩니다(라우터본 외. 1993,1995;파스쿠알 외. 1998;비존 외., 1999). 콜린 성 대뇌 피 질의 입력 및 대뇌 피 질의 소성의 메커니즘의 변조 기본 잠재적인 메커니즘을 식별 하기 위한 중요 한 피 질에서 세포를 생성 하는 핵의 식별. 현재 연구에서 우리는 글루타마틴성 뉴런에 의한 비교적 드문 생산과 함께 성인 피질에서 피질 가바알 성 세포의 주요 공급원으로 대뇌 피질 가바알 성 세포의 하위 집합을 확인합니다.
재료 및 방법
모든 피험자는 동물 보호를 위한 제도적 지침에 따라 치료되었다. 이전 연구에서 특정 세포 유형 소스의 식별을 금지 하는 대뇌 피 질의 응 고환 식 식의 낮은 내인성 수준으로 인해 우리는 로컬 콜 히 친 주입 함으로써 세포 체에서 전송 및 세포(슈베르트 외.)를 생산 하는 소마에서 검출 가능한 응 고환 항 원의 축적의 결과로 차단 하는 미세 소관 중 합,억제 수행. 1972;홋펠트 외. 1978 년,코너와 바론,1992). 이전 보고서에 따르면 콜히친 치료는 면역 조직 화학 기술을 사용하여 응급실의 검출을 증가시킵니다(코너 및 바론,1992). 또한,현재 연구에서 사용 된 것과 유사한 콜히친 수치는 뉴로 트로 핀 가족 구성원의 신경 분포에서 심한 이상을 유발하지 않습니다., 1997).조직 제제
8 명의 성인 피셔 344 쥐(수컷 4 마리,암컷 4 마리,할란 스프 라그 돌리)를 25 밀리그램/밀리그램 케타민,1.3 밀리그램/밀리그램 자일라진,및 0.25 밀리그램/밀리그램 아세 프로 마진 칵테일로 마취시키고,5 마리를 주사 하였다.0µl 콜히친(10µg/μl 에 aCSF)에 대뇌 위치는 아래에서 평가 0.5µl/min 를 사용하여 10µl 해밀턴 주사기입니다. 주입 후에,바늘은 4 분 동안 그 자리에 주위 조직에 충분한 유포를 허용하기 위하여 남아 있었습니다. 4 마리의 동물은 1 차 운동 피질(미디엄 1)에서+1.2 밀리미터 전방(에이/피)및 2.5 밀리미터 외측(미디엄/엘)에서 브레그마로 양측 주사를 받았다. 나머지 4 마리의 동물은 모두 전두엽 피질에서 일방적 인 주사를 받았다(에이/피:+3.(1)뇌척수액(1)뇌척수액(1)뇌척수액(1)뇌척수액(1)뇌척수액(1)뇌척수액(1)뇌척수액(1)뇌척수액(1)뇌척수액(1)뇌척수액(1)뇌척수액(1)뇌척수액(1)뇌척수액(1)뇌척수액(1)뇌척수액(1)뇌척수액(1)뇌척수액(1)뇌척수액(1)뇌척수액(1)뇌척수액(1)뇌척수액(1)뇌척수액(1)뇌척수액(1)뇌척수액(1)뇌척수액(1)뇌척수액(1)뇌척수액(1)뇌척수액( 250 밀리리터의 차가운 2%파라 포름 알데히드+0.2%파라 벤조 퀴논 0.1 엠 포스페이트 완충액. 뇌를 추출하고,동일한 정착액에서 2 시간 동안 후 고정하고,4 에서 적어도 72 시간 동안 30%수크로오스를 함유하는 0.1 미터 인산염 완충액에서 냉동 보호 하였다. 또한,냉동 슬라이딩 마이크로톰을 절단하여 면역조직화학을 위해 추가로 가공될 때까지 냉동보호제에 저장하였다.
이중 라벨 형광 면역 조직 화학
순차적 이중 라벨 면역 조직 화학을 사용하여 뉴런 발현을 시각화했습니다. 프리 플로팅 섹션을 트리스 완충 식염수로 세척하고,0.25%트리톤 엑스-100 으로 침투시킨 후,비특이적 라벨링을 5%당나귀 혈청으로 차단하였다. 1992 년,토끼 항체(코너와 바론)에서 1:1000 을 희석하여,트리톤 엑스 100 의 0.25%및 5%당나귀 혈청을 배양하였다. 1 차 항체 배양 후,당나귀 항 토끼 비오틴 공액 이그(1:200;벡터 연구소,벌링 게임,캘리포니아,미국)에서 섹션을 배양했습니다. 1:200 에서 3 시간 동안 1:200 으로 희석 한 후,1:200 에서 1:200 으로 희석 한 후,1:200 에서 1:200 으로 희석 한 후,1:200 에서 1:200 으로 희석 한 후,1:200 에서 1:200 으로 희석 한 후,1:200 에서 1:200 으로 희석 한 후,1:200 에서 1:200 으로 희석 한 후,1:200 에서 1:200 으로 희석 한 후,1:200 에서 1:200 으로 희석 한 후,1:200 에서 1:200 으로 2013 년 1 월 15 일,2013 년 1 월 15 일,2013 년 1 월 15 일,2013 년 1 월 15 일,2013 년 1 월 15 일,2013 년 1 월 15 일,2013 년 1 월 15 일,2013 년 1 월 15 일,2013 년 1 월 15 일,2013 년 1 월 15 일,2013 년 1 월 15 일,2013 년 1 월 15 일,2013 년 1 월 15 일,2013 년 1 월 15 일,2013 년(2015 년 12 월 1 일).(2015 년 12 월 1 일).(2015 년 12 월 1 일).(2015 년 12 월 1 일).(2015 년 12 월 1 일).(2015 년 12 월 1 일).(2015 년 12 월 1 일).(2015 년 12 월 1 일).(2015 년 12 월 1 일).(2015 년 12 월 1 일).(2015 년 12 월 1 일).(2015 년 12 월 1 일).(2015 년 12 월 2067919,1:1500; 최종적으로,섹션들을 세척하고,알렉사 플루오르 594 또는 488-공액 당나귀 안티마우스(인바이트로젠)에서 실온에서 3 시간 동안 배양하고,다시 세척하고,유리 슬라이드에 장착하고,플루오로마운트 지(미국 버밍핸 남부 생명공학)에서 커버슬립하였다. 가바알성 세포의 식별을 극대화하기 위해,가바알성 65 및 가바알성 67 항체를 공동-배양하였다. 섹션의 하위 집합은 커버슬립되었습니다 연장 금 와 다피(인 비트로 겐)세포핵의 시각화를 위해.
항체 특성화
본 연구에서 사용된 항체는 정제된 마우스에 대하여 토끼에서 제기된 친 화성-정제된 폴리클로날(코너 및 바론,1992)이다. 이 항체는 정제된 마우스 및 재조합 인간 핵분열체를 인식하지만,재조합 인간 핵분열체와 교차 반응하지 않는다(코너 및 바론,1996). 또한,쥐 뇌에서 면역 반응 패턴 발현 쥐 뇌(코너 및 바론,1997)에서 현장 분석에서 얻은 밀접 하 게 일치 합니다.
단클론 항체 갓-65(발달 연구 하이브리도마 뱅크,갓-6)는 쥐의 뇌로부터 정제된 갓 단백질 면역아피핀으로 쥐를 면역화함으로써 생성되었다. 쥐 뇌 균질화의 웨스턴 블롯 분석 결과,항체는 갓-65 를 선택적으로 인식하지만 갓-67 은 인식하지 않는 것으로 나타났다(장 및 고트립,1988). 추가 학문은 갓 6 항체가 갓 65 의 씨 말단의 아미노산 475-571 사이에서 있는 에피토프를 인식한ㄴ다는 것을 보여주었습니다(버틀러 외., 1993).
갓-67 마우스 모노클론(밀리포어,마브 5406,로트: 25010139)는 인간 갓-67 의 아미노산 잔기 4-101 에 대해 제기되었으며,쥐의 뇌(제조사의 기술 정보)의 웨스턴 블롯 분석에 대한 단일 67-고농도 대역을 인식한다. 항체를 뇌에서 면역 양성 신호가 없는 결과(이토 외., 2007).이 연구에서는 칼슘/칼모둘린 의존성 단백질 키나아제 2 의 알파 서브유니트를 구체적으로 인지하고 있다. 웨스턴 블롯 분석은 항체가 50 의 단일 밴드를 식별하고 인산화 된 형태와 비 인산화 된 형태를 모두 인식한다는 것을 보여줍니다(에론 두 및 케네디,1985).면역블로팅은 항체가 28-카빈다댄드를 인식하고,칼빈딘 디-9 케이,칼레티닌,미오신 경쇄 및 미오신 경쇄와 같은 유사한 분자와 반응하지 않는 것으로 나타났다.파발 부민(제조업체의 기술 정보). 칼빈딘을 이용한 전흡수는 병아리 및 쥐의 뇌에서 정제된 칼빈딘의 면역염색을 제거하는 것으로 나타났다(파스텔외., 1987).또한,개구리 근육에서 정제된 파발부민에 대해 면역화된 마우스로부터 수집되었다. 단클론 항체는 첫 번째 칼슘 2+-결합 부위에서 에피토프에 대해 지시되며 면역블롯 분석은 그것이 12 의 뇌 단백질을 인식한다는 것을 증명한다.
공초점 분석
이미지를 마그나파이어 소프트웨어와 함께 올림푸스 액스 70(버전 2.0;미국 캘리포니아 주 골레타 칼 스토츠 이미징)으로 촬영하였다. 코히친 확산의 부족과 체세포 코히친 축적의 결과 부재로 인해,각 코히친 주사 부위의 1.5 밀리미터 내의 섹션만 분석되었다. 피질 영역 내에서 10 번째 섹션(400,000,000)을 검사했습니다. 세포를 수동으로 계산 하 고 적어도 3 개의 섹션 각 피 질 영역 및 주제 당 항 체 조합에서 분석 했다. 만 3 동물 4 에서 감지할 수 없는 비단락 라벨링으로 인해 시간 영역에서 평가 되었다. 단일 및 이중 표지 셀 단일 및 이중 채널 이미지를 사용 하 여 정량화 했다. 다피 염색 조직을 평가할 때 크기,형태,신호 대 배경 및 일치 다피 라벨링을 포함하여 라벨링 된 세포를 식별하는 데 몇 가지 기준이 사용되었습니다. 에서 하드 디스크 드라이브에 저장 섹션,5μm z-stacks 수집되었을 사용하여 올림푸스 Fluoview FV1000 하도록 신경 지역화의 레이블이 있는 객체입니다. 비율,그리고 절대 수,이중 레이블된 셀의 우리의 관심으로 인해 입체 방법 사용 되지 않았습니다.
각 이미지 필드에 대해 면역 반응성(적외선)세포군 당 이중 표지 세포의 백분율을 결정하였다. 검사 된 각 피질 영역에 대해 평균 1000 표준 오차를 계산했습니다. 분산(분산 분석)의 단방향 분석은 피질 영역 간의 차이를 평가하는 데 사용되었습니다. 피셔의 고열장애는 사후 분석에 사용되었다. 모든 통계 분석은 윈도우 15.0 으로 수행되었다.
항체 대조군
대조군에는 1 차 항체의 생략,2 차 항체의 생략,및 1 차 항체를 비특이적 항체로 대체하는 것이 포함되었다. 모든 조작 예상된 효과 했다 하 고 그들의 해당 항 체에 의해 진정한 항 원 라벨 표시 셀 대표 하는 주장을 지원.
결과
면역 라벨링
구별된 면역 라벨링은 1 반경 내에서 볼 수 있었다.콜히친 피질 주사 부위로부터 5 밀리미터. 이 영역 내에서,세포 소마 타(그림 1)에 주로 국한되었습니다. 드물게 하나 이상의 세포 과정을 구별 할 수도 있습니다. 이 1.5 밀리미터 반경 밖에서는 피질에서 거의 감지 할 수 없었습니다. 으로 대뇌의 식 NGF 수용체 TrkA 및 p75 국 cholinergic corticopetal 섬유,관찰 라벨 NGF 가능성을 반영하여 endocytosed NGF 지만,대신 나타내는 NGF-생산 세포(holtzman 에 et al. 1995;로시 외.,2002;스티븐스 외., 2005). 이전에보고 된 바와 같이(리박등.,1978),콜히친 치료 신경 프로세스 뿐만 아니라 세포 시체에서 갓 라벨 강화. 콜히친은 파발 부민,칼빈 딘 또는 캄 키아에 대한 라벨링에 검출 가능한 효과가 없었다.
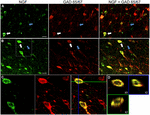
그림 1. 신경 성장 인자는 억제 세포 마커로 광범위하게 콜로코 화됩니다. 면역 반응 세포 에(ㅏ)일차 운동 피질 과(비)전두엽 피질. 이미지를 표시하는 NGF-labeled 세포(녹색)colocalize 으로 광범위하게 갓 65/67 라는 세포(적색). 흰색 화살표는 각 피질 영역에서 두 가지 모두에 대한 면역 반응성 세포의 예를 보여줍니다. 파란색 화살표는 갓 65/67 만 면역 반응 세포를 보여줍니다. (기음)에 흰색 상자 내부 영역의 확대 이미지(비). (디)엑스–와이–지 에 표시된 이중 표지 된 셀의 재구성(씨). 녹색 및 파란색 선은 각각 엑스 지 및 와이지 평면에서 해부 수준을 나타냅니다. 축척 막대=25,000,000,000.신경 성장 인자는 조사 된 피질 영역에 관계없이 가바 에르 세포 마커 가바 에르 65 및 가바 에르 67 과 광범위하게 공동 국소화되었다(그림 1;표 1). 피질세포의 91%는 피질세포의 9.9%에 해당하는 피질세포의 9.9%에 해당하는 피질세포의 9.9%에 해당하는 피질세포의 9.9%에 해당하는 피질세포이다. 의 백분율 NGF-labeled 세포 co-을 표현하는 GAD65/67 보여주는 작은 차이점은 전두엽(90.0±1.5%),모터(91.7±1.5%),정수리(89.6±3.5%)및 시간(93.4±8.1%)외피가(one-way ANOVA;p=0.78). 반대로,NGF co-localized55±2.모든 세포 중 3%가 표지 된 세포입니다. 가바 에르 뉴런 생성이 특정 하위 유형으로 제한되었는지 확인하기 위해,우리는 파발 부민 또는 칼빈딘(그림 2)에 대한 조직을 공동 표지했습니다. 두 마커와 함께 공동 화 하는 것이 관찰 되었다. 그러나,NGF colocalization parvalbumin(67.8±3.6%)었 2×이상 calbindin(29.1±3.9%). 또한,파발부민(47.7 4.6%)과 칼빈딘(25.7 4.9%)의 면역 반응성 세포의 절반 미만을 구성하였다.

그림 2. 억제 뉴런 서브 클래스 마커. 운동 피질의 조각들은 비엔티에프 및 파발부민 또는 칼빈딘에 대해 분류되었다. (에이)세포는 광범위한 중첩을 보였다. 1990 년대 초반,1990 년대 초반,1990 년대 초반,1990 년대 초반,1990 년대 초반. 축척 막대=25,000,000,000.

표 1. 피질 영역에 의한 면역 반응성 세포.
모든 피질 층에서 신경 성장 인자 발현 뉴런이 관찰되었다. 이전의 연구들은 대뇌 피질의 층류(피츠와 밀러,2000;파츠와 와일,2006)에서 비정형 뉴런 표지 뉴런의 분포가 불균일하다고 보고했다. 그러나 레이어에 의해 정량 분석,현재 연구에서 수행 되지 않았습니다.세포 소마타 및 근위 과정(그림 3)내에서 캄키아에 대한 라벨링이 주로 관찰되었다. (그림 3;표 2). 1.1%의 면역 반응성 세포를 캄 키아 항체와 공동 표지 하였다. 공동 현지화 대뇌 피 질 영역(단방향 분산 분석;피=0.03)에 의해 크게 달랐다 피셔의 포스트 특별 한 전 두 엽 피 질 기본 모터 피 질(2.4 1.0%;피=0.02)와 두 정 수 리 피 질(2.9%1.5;피=0.01)에 비해 이중 표지 세포(7.6 2.1%)의 더 큰 비율을 했다 밝혔다.

그림 3. 신경 성장 인자는 흥분성 세포 마커 캄 키아와 함께 최소한으로 공동 화됩니다. 전두엽 피질의 면역 반응 세포. 피질 영역을 조사한 것과 관계없이 세포는 녹색(녹색)과 캄 키아(빨간색)에 대해 거의 콜라 블링되지 않았습니다. 금 화살표는 면역 반응성 세포의 예를 보여줍니다. 축척 막대=25,000,000,000.

표 2. 피질 영역에 의한 면역 반응성 세포.
캄키아에 대한 면역반응성 세포는 항체에 의해 표지된 세포보다 훨씬 많다. 캄 키아 표지 된 세포의 전체 비율은 동시에 2,000,000,000 0.6%였다. 1 차 운동 피질(0.8%,피=0.001),정수리 피질(1.4%,피=0.01),측두엽 피질(1.5%,피=0.6%)보다 이중 표지 된 세포(3.7%,피 1.1%)의 더 큰 비율을 나타냈다.
토론
현재 연구는 피질의 핵피질 생성 뉴런의 대다수(>90%)가 가바 에르 인 반면,모든 가바 에르 뉴런 중 절반이 핵피질 세포로 공동 화된다는 것을 보여줍니다. 대조적으로,흥분성 뉴런의 마커는 희귀 한 공동 지역화를 나타냅니다. 이러한 결과이 연구에서 분석 하는 여러 대뇌 피 질 영역에 걸쳐 일관 되 게 했다.
비록 흥분성 세포 마커와 면역 반응성이 거의 일치하지는 않았지만,검사된 모든 피질 영역에 걸쳐 작은 비율(~5%)이 캄키아에 대해 양성이었다. 공 초점 분석 확인이 이중 라벨링 동일한 초점 평면에서 유래 하 고 중복 셀에서 발생 하는 개별 신호로 인해 아니었다. 흥분 성 세포의 이러한 작은 부분에서 알 수 없는 하지만 그것은 이러한 흥분 성 세포를 생산 하는 신경 피 질에서 흥분 성 뉴런의 이전에 알 수 없는 하위 클래스를 대표 하는 것이 가능 합니다.
우리의 결과는 다른 뇌 영역에서 초기 연구 결과와 일치하여 선조체,기저 전뇌 및 해마의 가바 성 세포와 거의 독점적으로 공동 국소화된다는 것을 보여줍니다(라우터본 외. 1993,1995;파스쿠알 외. 1998;비존 외., 1999). 따라서,가바 성 세포에 의한 응고막 생성은 기저 전뇌 콜린성 신경 분포(기저 전뇌 자체 포함)를 수신하는 모든 표적의 일반적인 특성 일 수있다. 이러한 콜린성 신경 세포 집단은 그들의 표현형 및 투영의 유지를 위해 비정형 신경 세포(라일 렛 외., 1993; 첸 등. 1997;후진타오 외. 1997;데베이르 외., 1999). 연구 전반에 걸쳐,억제 성 인터 뉴론은 이제 기저부 전뇌 콜린성 뉴런에 대한 핵 영양의 주요 원천으로 등장합니다.
비록 기저 전뇌 콜린성 뉴런이 흥분성 및 억제성 피질 네트워크 모두를 신경질적으로 자극하지만(자보르스키 외. 1999),영양 지원의 소스는 압도적으로 대뇌 피 질 회로(헨 쉬와 스트라이커,2004;야자 키-스기 야 마 등에서 소성을 매개에 중요 한 역할을 하는 것으로 알려져 억제 인구에서. 2009;도나 토 외., 2013). 콜린성 신호는 다양한 억제 하위 집단의 차동 활성화에 기여하여 흥분성 억제 균형을 조절하는 것으로 보입니다(시앙 외. 1998;프롬케 외., 2007). 이에 따라 고소 성 억제 네트워크와 이들을 활성화시키는 콜린성 입력 사이의 피드백 신호 역할을 할 수 있습니다. 콜린성 의존성 메커니즘을 통해 소성 및 행동 학습을 강화합니다.,1996,2004;구티에레즈 외. 1997;코너 외., 2009). 이 경우,이 약물은 다른 약물보다 더 많이 투여 될 수 있습니다. 2003),콜린성 활성을 증가시키기 위해 콜린성 프로바이오틱스(라일 렛 등)의 능력과 짝을 이루었다. 1993;후진타오 외. 1997;올드외.,2001),비정규직이 강화 된 콜린성 기능을 통해 활성 회로의 재구성을 촉진 할 수 있음을 시사한다.
향후 연구에서는 억제 뉴런의 어떤 하위 클래스가 비정형 뉴런을 발현하는지,이 표현이 피질 회로에 정확히 어떻게 영향을 미치는지 확인하려고 시도 할 것이다. 이를 위해,우리는 억제 뉴런,파발부민(주로 바구니와 샹들리에 세포와 관련된),그리고 칼빈딘(수많은 억제 세포 유형과 관련된 두 가지 일반적인 마커와 함께 지역화 여부를 결정하기 위해 예비 조치를 취했다;맥베인과 피산,2001;마크람 등., 2004). 우리의 결과 파 발 부 민 면역 반응 세포에서 주로 검출 되었다 비록 파 발 부 민에 의해 표시 하는 많은 셀 하지 않았다 공동 표현 시연. 중요 한 것은,억제 뉴런의 하나 이상의 하위 클래스를 생성 하는 것이 좋습니다.
요약하면,쥐의 신피질 내에서는 주로 억제 뉴런으로 표현되는데,이는 계통 발생 학적으로 오래된 뇌 영역에서 보존되고 전체적으로 뇌로 일반화 될 수있는 것으로 보인다. 우리의 결과 대뇌 피 질 억제 뉴런 기초 전뇌의 콜린 성 프로젝션 뉴런의 유지에 중요 한 역할을 하는 것이 좋습니다. 따라서 가바 에르 인터 뉴론은 규제 된 비정형 신호를 통해 피질 재구성을 촉진 할 수 있습니다.
이해 상충 성명
저자는 연구가 잠재적 인 이해 상충으로 해석 될 수있는 상업적 또는 재정적 관계가없는 상태에서 수행되었다고 선언합니다.
승인
에 의해 자금 지원 NIH(AG10435),재향 군인의 관리,알츠하이머의 협회 및 박사는 미리암과 Sheldon G. 아델슨 의료 연구 기초입니다.
보충 자료
이 기사의 보충 자료는 다음에서 온라인으로 찾을 수 있습니다: http://www.frontiersin.org/journal/10.3389/fncel.2014.00220/abstract
2010 년 11 월 15 일,2010 년 11 월 15 일,2010 년 11 월 15 일,2010 년 11 월 15 일,2010 년 11 월 15 일. 신경 성장 인자는 배양 된 기초 전뇌 뉴런에서 장기간 아세틸 콜린 방출을 빠르게 유도합니다:신경 조절 및 신경 영양 영향 사이의 분화. 신경 과학자. 21, 3375–3382.
2007 년 11 월 1 일. 기저 핵의 자극에 의해 대뇌 피질의 생리적 기억의 유도. 프록 내틀 아카드 과학. 미국 93,11219-11224. 도이:10.1073/나스.93.20.11219
교차 참조 전체 텍스트
2009 년 12 월 1 일,2009 년 12 월 1 일,2009 년 12 월 1 일,2009 년 12 월 1 일,2009 년 12 월 1 일. 선조체 인터 뉴론의 하위 집단은 신경 영양 인자 발현에 기초하여 구별 될 수있다. 제이 컴포지션. 뉴롤이야 408, 283–298. 도이:10.1002/(시치)1096-9861(19990531)408:2<283::5547>3.0.CO;2-2
펍메드 초록/펍메드 전체 텍스트/크로스레프 전체 텍스트
2010 년 11 월 15 일. 해 마 뉴런에서 신경 성장 인자 릴리스의 특성:구성 및 틀에 얽매이지 않는 나트륨 종속 규제 경로에 대 한 증거. 유로 신경 과학자. 7, 1220–1228. 도이:10.1111/제이.1460-9568.1995.01112.전체 텍스트|교차 참조 전체 텍스트|초록/초록/초록/초록/초록/초록/초록/초록/초록/초록/초록/초록/초록/초록/초록/초록
2009 년 12 월 15 일,2009 년 12 월 15 일,2009 년 12 월 15 일,2009 년 12 월 15 일,2009 년 12 월 15 일,2009 년 12 월 15 일,2009 년 12 월 15 일. 글루탐산 탈 카르 복실 라제의 지배적 인 에피토프의 식별(갓-65)뻣뻣한 남자 증후군에서자가 항체에 의해 인식. 제 1 조(특급) 메드. 178, 2097–2106. 도이:10.1084/젬.178.6.2097
펍메드 초록/펍메드 전체 텍스트/크로스레프 전체 텍스트
2010 년 11 월 1 일,2010 년 11 월 1 일. 글루탐산 탈 카르 복실 라제에 대한 단일 클론 항체로 정제 된 단백질의 특성. 신경 과학자. 8, 2123–2130.
2009 년 12 월 1 일,2009 년 12 월 1 일,2009 년 12 월 1 일,2009 년 12 월 1 일,2009 년 12 월 1 일,2009 년 12 월 1 일,2009 년 12 월 1 일,2009 년 12 월 1 일. 신경 성장 인자 유전자의 단일 대립 유전자의 붕괴는 기초 전뇌 콜린성 뉴런 및 기억 결핍의 위축을 초래합니다. 신경 과학자. 17, 7288–7296.
2005 년 12 월 15 일,2005 년 12 월 15 일,2005 년 12 월 15 일,2005 년 12 월 15 일,2005 년 12 월 15 일. 기초 전뇌 콜린성 시스템은 뇌 손상 후 대뇌 피질의 가소성 및 기능 회복에 필수적입니다. 뉴런 46,173-179. 도이:10.1016/제이.2005.03.003
펍메드 초록/펍메드 전체 텍스트/크로스레프 전체 텍스트
코너,제이 엠.,Culberson,A.,Packowski,C.,치바,A.,및 Tuszynski,M.H.(2003). 기초 전뇌 콜린성 시스템의 병변은 작업 획득을 손상시키고 운동 기술 학습과 관련된 대뇌 피질의 가소성을 폐지합니다. 뉴런 38,819-829. doi:10.1016/S0896-6273(03)00288-5
Pubmed 추상|Pubmed 전체 텍스트|CrossRef 전체 텍스트
코너,J.M.,프랭크,K.M.,Titterness,A.K.,러셀,K.,Merrill,D.A.,크리스티,R.B.,et al. (2009). 해마 소성 및 학습에 필수적입니다. 신경 과학자. 29, 10883–10889. 도이:10.1523/뉴로시.2594-09.2009 년
Pubmed 추상|Pubmed 전체 텍스트|CrossRef 전체 텍스트
코너,J.M.,Lauterborn,J.C.,연,Q.,담낭,C.M.,과오,S.(1997). 성인 쥐의 중추 신경계에서 뇌 유래 신경 영양 인자의 분포:전 행성 축삭 수송에 대한 증거. 신경 과학자. 17, 2295–2313.
2009 년 11 월 1 일. 콜히친 치료 후 성인 쥐 뇌에서 신경 성장 인자와 같은 면역 반응 뉴런의 분포. 제이 컴포지션. 뉴롤이야 326, 347–362. 도이:10.1002/중국 중부 표준시.903260304
펍메드 초록/펍메드 전체 텍스트/크로스레프 전체 텍스트
2009 년 12 월 15 일. 신경 성장 인자에 대한 항체의 특성화:다른 뉴로 트로핀과의 교차 반응성의 분석 의존적 변동성. 신경 과학자. 방법 65,93-99. 도이: 10.1016/0165-0270(95)00151-4
펍메드 초록/펍메드 전체 텍스트/크로스레프 전체 텍스트
2009 년 11 월 1 일. 쥐의 뇌에서 면역 반응성의 발달 프로필: 해 마와 피 질에서 콜린 성 터미널 필드의 설립에 핵의 가능한 역할. 뇌 레스. 뇌 재.101,67-79. 10.1016/0165-3806(97)00051-5
펍메드 초록/펍메드 전체 텍스트/크로스레프 전체 텍스트
2010 년 12 월 1 일,2010 년 12 월 1 일,2010 년 12 월 1 일,2010 년 12 월 1 일,2010 년 12 월 1 일,2010 년 12 월 1 일. 신경 성장 인자 모방 트라 카 길항제는 성인 쥐에서 피질 콜린성 부톤의 철수를 유발합니다. 프록 내틀 아카드 과학. 미국 96,4067-4072. 도이:10.1073/나스.96.7.4067
펍메드 초록/펍메드 전체 텍스트|크로스레프 전체 텍스트
2013 년 11 월 15 일,2013 년 11 월 15 일,2013 년 11 월 15 일,2013 년 11 월 15 일. 경험에 의해 유도 된 파발 부민 표현 바구니 세포 네트워크 가소성은 성인 학습을 조절합니다. 자연 504,272-276. 10.1038/자연 12866
펍메드 초록|펍메드 전체 텍스트/크로스레프 전체 텍스트
2010 년 11 월 1 일. 쥐 뇌에서 제 2 형 카 2+/칼모 둘린 의존성 단백질 키나아제의 지역 분포. 신경 과학자. 5, 3270–3277.
프롬케,아르 자형. 2007 년 11 월 15 일,2007 년 11 월 15 일,2007 년 11 월 15 일,2007 년 11 월 15 일. 대뇌 피 질의 수용 필드 소성에 대 한 시 냅 스 메모리 추적. 자연 450,425-429. 10.1038/네이처 06289
펍메드 초록/펍메드 전체 텍스트/크로스레프 전체 텍스트
2010 년 11 월 1 일. 변연 발작은 신경 성장 인자에 대한 메신저 뉴런 생성을 증가시킵니다. 과학 245,758-761. 도이:10.1126/과학.2549634
교차 참조 전체 텍스트
2009 년 12 월 1 일,2009 년 12 월 1 일,2009 년 12 월 1 일,2009 년 12 월 1 일,2009 년 12 월 1 일,2009 년 12 월 1 일. 피질 신경 성장 인자 박탈 후 학습 장애 및 콜린성 탈진. 신경 과학자. 17, 3796–3803.
Pubmed 추상|Pubmed 전체 텍스트
Hanson,M.,and Edström,A.(1978). 유사 분열 억제제 및 축삭 수송. 지능. 사이톨 목사. 지원 373–402.
2009 년 12 월 1 일,2009 년 12 월 1 일,2009 년 12 월 1 일,2009 년 12 월 1 일,2009 년 12 월 1 일,2009 년 12 월 1 일,2009 년 12 월 1 일. 원숭이중추신경계에서 신경성장인자에 대한 유전자의 발현. 뇌 618,142-148. 도이:10.1016/0006-8993(93)90437-아르 자형
교차 참조 전체 텍스트
1986. 신경 성장 인자는 중격 절곡 후 중격 콜린성 뉴런의 생존을 촉진합니다. 신경 과학자. 6, 2155–2162.
2010 년 11 월 1 일. 고양이 시각 피질을 개발하는 가바 회로에 의해 조각 된 원주 형 건축. 과학 303,1678-1681. 도이:10.1126/과학.1091031
펍메드 초록/펍메드 전체 텍스트/크로스레프 전체 텍스트
2015 년 11 월 1 일,2015 년 11 월 1 일,2015 년 11 월 1 일. 고양이 1 차 감각 뉴런에서 물질의 국소화 및 분포에 대한 실험적 면역 조직 화학적 연구. 뇌 재.100,235-252. doi: 10.1016/0006-8993(75)90481-3
Pubmed 추상|Pubmed 전체 텍스트|CrossRef 전체 텍스트
holtzman 에,D.M.,Kilbridge,J.,Li,Y.,커닝햄,E.T.,Jr.,Lenn,N.J.,클러리,D.O.,et al. (1995). 중추신경계의 트라카 표현:몇몇 신규한 중추신경 신경세포의 존재에 대한 증거. 신경 과학자. 15, 1567–1576.
후,엘. 2010 년 11 월 1 일. 핵의 콜린성 표현형의 차동 변조 기저 세포 신경 세포 세포 또는 대뇌 피질 말단 필드에. 특급. 뉴롤이야 143, 162–171. 도이:10.1006/외출수.1996.6357
펍메드 초록/펍메드 전체 텍스트/크로스레프 전체 텍스트
2010 년 11 월 15 일,2010 년 11 월 15 일. 신경 세포 수용체:신경 세포 신호 전달의 역할. 아누 바이오켐 목사. 72, 609–642. 도이:10.1146/아누 레프.바이오켐72.121801.161629
교차 참조 전체 텍스트
2009 년 12 월 1 일,2009 년 12 월 1 일,2009 년 12 월 1 일,2009 년 12 월 1 일,2009 년 12 월 1 일,2009 년 12 월 1 일,2009 년 12 월 1 일. “………..”………..”………..”. 뉴런 6,937-948. 10.1016/0896-6273(91)90234-큐
펍메드 초록/펍메드 전체 텍스트/크로스레프 전체 텍스트
이토,티,히오키,에이치,나카무라,케이,다나카,와이이,나카데,에이치,가네코,티 등이 있다. (2007). 쥐 흉부 척수의 감마-아미노 부티르산 함유 교감 신경절 전 뉴런은 축삭을 상 경부 신경절로 보냅니다. 제이 컴포지션. 뉴롤이야 502, 113–125. 도이:10.1002/중국 중부 표준시.21309
펍메드 초록/펍메드 전체 텍스트/크로스레프 전체 텍스트
2009 년 12 월 1 일,2009 년 12 월 1 일,2009 년 12 월 1 일,2009 년 12 월 1 일,2009 년 12 월 1 일. 핵 기저 활동에 의해 활성화 된 피질지도 재구성. 과학 279,1714-1718. 도이:10.1126/과학.279.5357.1714
펍메드 초록/펍메드 전체 텍스트/크로스레프 전체 텍스트
코더워,제이,바투스,알티 2009 년 12 월 1 일,2009 년 12 월 1 일,2009 년 12 월 1 일,2009 년 12 월 1 일,2009 년 12 월 1 일. 비인간 영장류의 신경 성장 인자 수용체 면역 활동(세 버스 아 펠라):콜린성 효소를 이용한 분포,형태 및 대장 화. 제이 컴포지션. 뉴롤이야 277, 465–486. 도이:10.1002/중국 중부 표준시.902770402
펍메드 초록/펍메드 전체 텍스트/크로스레프 전체 텍스트
100,000,000,000,000,000,000,000,000,000,000,000,000,000,000,000,000,000,000,000,000,000,000,000,000,000,000,000,000 (1994). 나이 든 원숭이 기초 전뇌: 인간 신경 성장 인자를 분비하는 캡슐화 된 세포의 이식 후 축 절제 된 기초 전뇌 뉴런의 구조 및 발아. 프록 내틀 아카드 과학. 미국 91,10898-10902. 도이:10.1073/나스.91.23.10898
Pubmed 추상|Pubmed 전체 텍스트|CrossRef 전체 텍스트
Korsching,S.,Auburger,G.,Heumann,R.Scott,J.,그리고 Thoenen,H.(1985). 쥐의 중추 신경계에 신경 성장 인자와 그 미나의 수준은 콜린성 신경 분포와 상관 관계. 4,1389-1393.
2010 년 11 월 15 일-2010 년 11 월 15 일. 신경 성장 인자:알츠하이머 병에 대한 구조,기능 및 치료 적 영향. 커 약물은 중추 신경계 뉴롤을 표적으로합니다. 불화. 2, 315–334. doi: 10.2174/1568007033482724
Pubmed 추상|Pubmed 전체 텍스트|CrossRef 전체 텍스트
큰,T.H.,Bodary,S.C.,클레,D.O.,Weskamp,G.,오튼,미국,그리고 Reichardt,L.F.(1986). 개발 쥐 뇌의 신경 성장 인자 유전자 발현. 과학 234,352-355. 도이:10.1126/과학.3764415
교차 참조 전체 텍스트
2009 년 12 월 1 일,2009 년 12 월 1 일,2009 년 12 월 1 일,2009 년 12 월 1 일,2009 년 12 월 1 일,2009 년 12 월 1 일,2009 년 12 월 1 일. 쥐의 기저부 전뇌에서 콜린성 뉴런이 아닌 가바 성 뉴런에 의해 표현된다. 제이 컴포지션. 뉴롤이야 360, 454–462. 도이:10.1002/중국 중부 표준시.903600307
펍메드 초록/펍메드 전체 텍스트/크로스레프 전체 텍스트
2009 년 12 월 1 일,2009 년 12 월 1 일,2009 년 12 월 1 일,2009 년 12 월 1 일,2009 년 12 월 1 일,2009 년 12 월 1 일,2009 년 12 월 1 일. 신경 성장 인자는 쥐 해마에서 가바 성 뉴런에 의해 표현됩니다. 뉴로 포트 5,273-276. 도이:10.1097/00001756-199312000-00023
크로스레프 전체 텍스트
2010 년 11 월 15 일,2010 년 11 월 15 일,2010 년 11 월 15 일,2010 년 11 월 15 일. 뉴로 트로 핀 분비:현재 사실과 미래 전망. 음식물. 뉴로바이올 69, 341–374. doi:10.1016/S0301-0082(03)00019-4
Pubmed 추상|Pubmed 전체 텍스트|CrossRef 전체 텍스트
Li,Y.,holtzman 에,D.M.,Kromer,L.F.,Kaplan,D.R.,추아-Couzens,J.,클러리,D.O.,et al. (1995). 랫 기초 전뇌 개발 시 트르카 및 채팅 표현 조절: 콜린성 뉴런의 분화를 조절한다는 증거. 신경 과학자. 15, 2888–2905.
이 경우,본 발명의 다른 실시예들은,예를 들어,본 발명의 다른 실시예들로서,상기 제 1 실시예들,또는 제 2 실시예들,및 제 2 실시예들,및 제 2 실시예들,및 제 2 실시예들,및 제 2 실시예들,및 제 2 실시예들,및 제 2 실시예들,및 제 2 실시예들,및 제 2 실시예들,및 제 2 실시예들,및 제 2 실시예들,및 제 2 실시예들,및 (1990). 쥐의 신경계에는 평행 및 상호 표현 패턴이 있다. 뉴런 5,501-509. 쩌짯쩔챦짱쨘째쩍 짹쨍쨌짹쨍 쨉챨철쨌 32 짹챈 30,13 첸 쩔짙쩔징징쩔짙째챠쨉쨉()째챠째쨈쩌쩐(1688-7662)째챠째쨈쨈 쨔짰쨩챌쨘(1688-7662)쨌 쩍쩍쩔
마크 람,에이치. 톨레도-로드 리 게스,M.,Wang,Y.,Gupta,A.,다,G.,and Wu,C.(2004). 신피질 억제 시스템의 인터 뉴런. 냇 신경 과학자. 5, 793–807. 10.1038|1519
10.1038|1519
10.1038/151910.1038/151910.1038
2010 년 11 월 1 일. 인터 뉴런 언 바운드. 냇 신경 과학자. 2, 11–23. doi: 10.1038/35049047
Pubmed 추상|Pubmed 전체 텍스트|CrossRef 전체 텍스트
Mufson,E.J.,코너,J.M.,오,S.,and Kordower,J.H.(1994)등이 있습니다. 영장류 기저부 전뇌 및 해마 형성에서 신경 성장 인자와 유사한 면역 반응 프로파일. 제이 컴포지션. 뉴롤이야 341, 507–519. 도이:10.1002/중국 중부 표준시.903410407
펍메드 초록/펍메드 전체 텍스트/크로스레프 전체 텍스트
2009 년 12 월 1 일,2009 년 12 월 1 일,2009 년 12 월 1 일,2009 년 12 월 1 일,2009 년 12 월 1 일,2009 년 12 월 1 일,2009 년 12 월 1 일. 해마 인터 뉴런에서의 신경 성장 인자 및 뉴로 트로 핀-3 의 발현:형태 학적 특성,발현 수준 및 신경 성장 인자 및 뉴로 트로 핀-3 의 공동 화. 제이 컴포지션. 뉴롤이야 395, 73–90. 도이:10.1002/)1096-9861(19980525)395:1<73::5547>3.0.CO;2-전자
펍메드 초록/펍메드 전체 텍스트/크로스레프 전체 텍스트
2011 년 12 월 1 일,2012 년 12 월 1 일,2013 년 12 월 1 일,2013 년 12 월 1 일,2013 년 12 월 1 일,2013 년 12 월 1 일. 또한,면역 조직 화학적 교차 반응성 및 전기 영동 코 마이 그 레이션,칼빈딘 및 비시 닌 사이의 상호 반응성 및 전기 영동 코 마이 그 레이션. 412,107-113. 도이: 10.1016/0006-8993(87)91444-2
펍메드 초록/펍메드 전체 텍스트/크로스레프 전체 텍스트
2006. 쥐 시각 피질의 층에서 뉴로 트로 핀 미르 나 발현의 발달 변화. 유로 신경 과학자. 24, 2453–2460. 도이:10.1111/제이.1460-9568.2006.05126.전체 텍스트|교차 참조 전체 텍스트|초록/초록/초록/초록/초록/초록/초록/초록/초록/초록/초록/초록/초록/초록/초록/초록
2000 년 10 월 15 일.2000 년 10 월 15 일. 성숙한 쥐의 체성 감각 피질에서 신경 성장 인자,뇌 유래 신경 영양 인자 및 뉴로 트로 핀-3 의 발현:고 친 화성 뉴로 트로 핀 수용체와의 공동 표현. 제이 컴포지션. 뉴롤이야 418, 241–254. 도이:10.1002/(시치)1096-9861(20000313)418:3<241::1>3.0.CO;2-엠
펍메드 초록/펍메드 전체 텍스트|크로스레프 전체 텍스트
1996 년 12 월 19 일,1996 년 12 월 19 일,1996 년 12 월 19 일,1996 년 12 월 19 일,1996 년 12 월 19 일. 생체 내 성인 피질의 기능적 조직에 대한 비단배엽 피질 및 비단배엽 피질의 신속하고 반대 효과. 자연 381,702-706. 10.1038/381702 10
10.1038|10.1038|10.1038/10.1038/10.1038/10.1038/10.1038/10.1038
2010 년 11 월 15 일,2010 년 11 월 15 일,2010 년 11 월 15 일,2010 년 11 월 15 일,2010 년 11 월 15 일. 기저 전뇌 콜린성 시스템은 성인 쥐의 배럴 피질에서 빠른 신경 성장 인자(에프)에 의한 가소성에 관여한다. 제이.뉴로피시올. 91, 424–437. 도이:10.1152/요.00489.2003
펍메드 초록/펍메드 전체 텍스트/크로스레프 전체 텍스트
2011 년 12 월 1 일,2012 년 12 월 1 일,2013 년 12 월 1 일,2013 년 12 월 1 일,2013 년 12 월 1 일. 축삭 수송의 콜히친 억제에 따른 신경 소마타에서 글루탐산 탈 카르 복실 라제의 면역 세포 화학적 국소화. 뇌 재.140,315-332. 도이:10.1016/0006-8993(78)90463-8
펍메드 초록/펍메드 전체 텍스트/크로스레프 전체 텍스트
2010 년 11 월 15 일,2010 년 11 월 15 일. 쥐의 시각 피질에서 신경 성장 인자 수용체의 발현:콜린성 입력에 의한 발달 및 조절. 신경 과학자. 22, 912–919.
1993 년 12 월 15 일. 성인 및 노인 피셔 -344 쥐의 뇌내 투여 후 아세틸 콜린 합성 및 방출. 신경 과학자. 13, 3956–3963.
1972 년 12 월 15 일,1972 년 12 월 15 일,1972 년 12 월 15 일,1972 년 12 월 15 일,1972 년 12 월 15 일. 수상 돌기에서의 신경 플라스마 수송:고양이의 운동 신경 세포의 형태 및 생리학에 대한 콜히친의 효과. 뇌 레스 47,331-343. 도이: 10.1016/0006-8993(72)90643-9
펍메드 초록/펍메드 전체 텍스트/크로스레프 전체 텍스트
2018 년 11 월 1 일 (1986). 베타 신경 성장 인자(중추신경계)유전자의 발현에 관한 연구: 프록 내틀 아카드 과학. 미국 83,2714-2718. 도이:10.1073/나스.83.8.2714
펍메드 초록/펍메드 전체 텍스트/크로스레프 전체 텍스트
2010 년 11 월 15 일,2010 년 11 월 15 일,2010 년 11 월 15 일,2010 년 11 월 15 일,2010 년 11 월 15 일. 신경 성장 인자 신호,신경 보호 및 신경 복구. 아누 신경 과학자. 24, 1217–1281. 도이:10.1146/아누 레프.신경외과24.1.1217
CrossRef 전체 텍스트
스티븐스,H.E.,Belliveau,A.C.,Gupta,J.S.,Mirkovic,S.,and Kablar,B.(2005). 척수 운동 뉴런 및 등쪽 뿌리 신경절 고유 감각 감각 뉴런의 유지에서 뉴로 트로핀의 역할. 지능. 제이 데브. 신경과학자 23, 613–620. 도이:10.1016/제이.2005.07.002
펍메드 초록/펍메드 전체 텍스트/크로스레프 전체 텍스트
2010 년 11 월 15 일,2010 년 11 월 15 일,2010 년 11 월 15 일,2010 년 11 월 15 일,2010 년 11 월 15 일. 핌브리아/포닉스 절단에 따른 기저 전뇌 세포 손실. 뇌 재. 508, 241–248. 10.1016/0006-8993(90)90402-
교차 참조 전체 텍스트
2009 년 12 월 1 일,2009 년 12 월 1 일,2009 년 12 월 1 일,2009 년 12 월 1 일,2009 년 12 월 1 일. 신피질 억제 네트워크 내의 콜린성 전환. 과학 281,985-988. 도이:10.1126/과학.281.5379.985
펍메드 초록/펍메드 전체 텍스트/크로스레프 전체 텍스트
2009 년 12 월 15 일,2009 년 12 월 15 일,2009 년 12 월 15 일,2009 년 12 월 15 일,2009 년 12 월 15 일,2009 년 12 월 15 일,2009 년 12 월 15 일. 시각적 경험에 의한 빠른 스파이크 가바 회로에서 양방향 가소성. 자연 462,218-221. 도이:10.1038/네이처 08485
펍메드 초록/펍메드 전체 텍스트/크로스레프 전체 텍스트
2011 년 12 월 1 일,2012 년 12 월 1 일,2013 년 12 월 1 일,2013 년 12 월 1 일,2013 년 12 월 1 일,2013 년 12 월 1 일,2013 년 12 월 1 일,2013 년 12 월 1 일. 기저 전뇌 대뇌 피질 시스템을 재검토했습니다. 앤 뉴욕. 과학. 877, 339–367. 도이:10.1111/제이.1749-6632.1999.09276.x
Pubmed 추상|Pubmed 전체 텍스트|CrossRef 전체 텍스트
장,H.T.,리,L.Y.,Zou,X.L.,Song,X.B.,Hu,Y.L.,Feng,Z,T.,et al. (2007). 면역조직화학적 분포는 성인 히말라야 원숭이의 뇌에서 발견된다. J. Histochem. Cytochem. 55, 1–19. doi: 10.1369/jhc.6A6952.2006
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text