Introducere
ciclul glioxilat ocolește etapele de decarboxilare ale ciclului Krebs și determină asimilarea carbonului lichid din acetil-CoA. Izocitrat liaza (EC 4.1.3.1) și malat sintaza (EC 2.3.3.9) sunt singurele enzime pentru această cale metabolică. Ciclul glioxilat are loc în germinarea semințelor (Eastmond & Graham, 2001), la unele nevertebrate, cum ar fi C. elegans (Edwards, Copes, Brito, Canfield, & Bradshaw, 2013) și în microorganisme, cum ar fi E. coli (Nelson & Cox, 2009) și S. cerevisiae (Rezaei, Aslankoohi, Verstrepen, & Courtin, 2015). Calea a fost descrisă în 1957 în timpul studiilor asupra microorganismelor care au crescut pe acetat și etanol ca sursă de carbon (Kornberg & Krebs, 1957).
acetil-CoA în ciclul glioxilat se leagă de oxaloacetat, respectiv producând citrat și izocitrat. Acesta din urmă produce glioxilat și succinat în reacția catalizată de izocitrat liază. Mai mult, glioxilatul se condensează cu o a doua moleculă de acetil-CoA într-o reacție catalizată de malat sintază, după ce malatul produs este oxidat la oxaloacetat și astfel regenerând intermediarul (etapele B-1 până la B-5, Figura 1). Două molecule de acetil-CoA sunt introduse la fiecare repetare a ciclului glioxilat și are loc sinteza lichidă a unei molecule de succinat (Beeckmans, 2009; Berg, Stryer, & Tymoczko, 2015; Torres & Marzzoco, 2007).
mai mult, carbohidrații produși din lipidele de rezervă din semințele oleaginoase sunt distribuite plantei și utilizate ca sursă de carbon până când cloroplastele încep să se diferențieze în folioles și încep fotosinteza. Grăsimile și uleiurile sunt structuri chimice importante în stocarea carbonului multor semințe, deși într-o formă redusă, cuprinzând specii agronomice importante precum soia, arahidele, bumbacul și floarea-soarelui (Junqueira, 2012). Acizii Lauric (12:0), myristic (14:0), palmitic (16:0) și stearic (18:0) sunt cei mai importanți acizi grași saturați din legume; în timp ce acizii nesaturați sunt acidul oleic [18:1 (Inqux9)], acidul linoleic [18:2(Inqux9, 12)] și acidul linolenic [18:3 (Inqux9, 12, 15)] (Taiz & Zeiger, 2002).

Figura 1.
acizii grasi sau catabolismul acetat produc energie prin ciclul glioxilat, evidentiind integrarea intre oxidarea-oxidata(etapele a-1 pana la a-4), ciclul glioxilat (etapele B-1 pana la B-5), ciclul Krebs (etapele C-1 pana la C-8), gluconeogeneza (etapele D-1 pana la D-3) si glicoliza (etapa E-1).
acizii grași cu un număr de perechi de atomi de carbon nu au ca rezultat sinteza carbohidraților la animale (Nelson & Cox, 2009). Acest fapt apare deoarece nu există o producție netă de oxaloacetat care să susțină gluconeogeneza, deoarece, în acest caz, acizii grași sunt catabolizați în acetil-CoA. Intermediarul este oxidat în ciclul Krebs și pentru fiecare doi atomi de carbon care intră sub formă de acetil-CoA, doi atomi de carbon se pierd sub formă de CO2 (Eastmond & Graham, 2001; Nelson & Cox, 2009).
absența ciclului glioxilat la mamifere poate fi o țintă a toxicității selective în dezvoltarea agenților antimicrobieni. Activitatea inhibitoare a 3-nitropropionamidelor pe izocitrat lyase în M. tuberculosis (Sriram și colab., 2011), bromofenoli (Oh și colab., 2010) și alcaloizi din bureți pe izocitrat lyase în C. albicans (Lee și colab., 2008; Oh și colab., 2010) au fost analizate.
nu există date în literatură cu privire la aspectele energetice care implică catabolismul lipidic prin ciclul glioxilat. În acest context, cercetarea de față a investigat relația dintre cantitatea de energie produsă în ciclul glioxilat, cu dimensiunea lanțului de acizi grași, și a comparat eficiența energetică a celulelor care utilizează această abordare, cu celulele animale lipsite de ciclul glioxilat.
Materialși metode
s-a realizat un echilibru energetic al etapelor cu producția și consumul de energie pentru a obține o ecuație care corelează direct dimensiunea lanțului unui acid gras cu un număr de carbon multiplu de 4, cu energia produsă prin degradarea acestuia. În ecuația generată, costul energetic implicat în activarea acidului gras pentru acil-CoA nu a fost contabilizat. Acesta este un pas înainte de oxidarea cu un nivel ridicat de energie și implică ruperea a două legături fosfatice de mare energie.
ecuația propusă a fost validată prin compararea cantității ATP obținute prin aplicarea sa, cu ratele ATP obținute prin adăugarea cantității ATP produse prin fosforilarea oxidativă și fosforilarea nivelului substratului în fiecare etapă a căilor metabolice care participă la degradarea acizilor grași care implică ciclul glioxilat. Degradarea acizilor grași implică oxidarea inqq în glioxizomi, ciclul glioxilat, ciclul Krebs, parte a gluconeogenezei și glicolizei, decarboxilarea piruvatului și din nou ciclul Krebs (Figura 1).
eficiența energetică a degradării acizilor grași care implică ciclul glioxilat a fost comparată cu catabolismul acizilor grași din celulele animale. În această comparație s-au folosit ecuații validate anterior și s-a calculat cantitatea de energie produsă în oxidarea de la X-X așa cum se întâmplă la animale (Gon-X-X, Valduga, & Pereira, 2012), cuplată la ecuațiile dezvoltate în cercetările actuale. Factorul de eficiență energetică în ciclul glioxilat (e%) a fost definit de raportul dintre ATP produs în catabolismul acizilor grași, luând în considerare ciclul glioxilat și catabolismul animal. Parametrul a fost reprezentat în funcție de numărul de atomi de carbon ai acidului gras. Mai mult, rata ATP produsă de unitatea CH2 a fost reprezentată în funcție de numărul de atomi de carbon ai lanțului de acizi grași. GrapPad Prism 6.0 a generat modelele matematice care au reprezentat relația dintre variabile.
Rezultateși discuție
matricea mitocondrială este locul principal în care are loc oxidarea acizilor grași în celulele animale. Cu toate acestea, catabolismul acestor biomolecule apare în principal în peroxizomii țesuturilor foliare și în glioxizomi în timpul germinării semințelor plantelor (Buchanan, Gruissem, Vickers, & Jones, 2015). Degradarea acizilor grași în glioxizomi include căile metabolice prezentate în tabelul 1.
în paralel cu producția de energie, conversia acizilor grași în carbohidrați are alte funcții, cum ar fi producerea de polizaharide structurale și nucleotide prin calea fosfatului de pentoză (Nelson & Cox, 2009). În ceea ce privește câștigul de energie, calea mai scurtă și mai economică din punct de vedere energetic, probabil selectată prin evoluție, include gluconeogeneza până la fosfoenolpiruvat (etapa D-3 din Figura 1), degradată pe calea glicolitică.
Cantitatea de energie (ATP) produsă în timpul degradării glioxizomale a acizilor grași
cantitatea de ATP (x) rezultată din degradarea glioxizomală a acizilor grași cu numărul de carbon multiplu de 4, urmată de gluconeogeneză până la fosfoenolpiruvat și catabolismul acestuia, pentru a produce energie,poate fi calculată prin suma ratelor de ATP produse și consumate în oxidarea glyox), ciclul Krebs (ATP c Krebs),gluconeogeneza (ATP oxal-PEP) și din nou ciclul glicolizei /Krebs (ATP pep-CO2), conform ecuației (1):

fiecare ruptură a legăturii carbon-carbon în timpul oxidării cu XIX în glioxizomi și peroxizomi produce un NADH care, fosforilarea oxidativă, produce 2,5 ATPs (etapa a-3 din Figura 1). În peroxizomi, energia electronilor eliminată în prima etapă a oxidării cu oxid de oxid este disipată sub formă de căldură (stepA-1 în Figura 1).

prin urmare, nu există energie netă din oxidarea FADH2 (Buchanan și colab., 2015). Din moment ce carbonul de la hectolitru este esterificat cu coenzima A, oxidarea de la hectolitru are loc 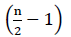 ori, unde n este numărul de carbon din lanțul de acizi grași. Cantitatea de ATPs produsă prin oxidarea în ciclul glioxilat este dată prin ecuație (2):
ori, unde n este numărul de carbon din lanțul de acizi grași. Cantitatea de ATPs produsă prin oxidarea în ciclul glioxilat este dată prin ecuație (2):

pentru fiecare două molecule de acetil-CoA direcționată către ciclul glioxilat, există o producție netă de o succinatmoleculă, care corespunde la jumătate din cantitatea de acetil-CoA, astfel încât moleculele de n/4succinat sunt produse. În ciclul Krebs, succinatul este oxidat la oxaloacetat, producând 1 FADH2 și 1 NADH care produce 4 ATP în fosforilarea oxidativă (etapele C-6 și C-8, figura 1). Această valoare înmulțită cu moleculele de succinat n/4 este exprimată prin ecuația (3), care reprezintă ATPamount produs mai întâi în ciclul Krebs:

regenerarea ciclului oxalacetat inglioxilat implică oxidarea malatului, care produce 1 NADH care produce 2,5 ATPs în fosforilarea oxidativă (etapa B-5 din Figura 1). În consecință, cantitatea de energie (ATP) produsă în ciclul glioxilat poate fi exprimată prin ecuație(4):

excesul de oxaloacetat produs în Krebscycle poate fi direcționat către gluconeogeneză. Pentru a produce energie, gliconeogeneza are loc în formarea fosfoenolpiruvatului, cu 1 GTP cheltuit (stepD3 în Figura 1), exprimat în ecuația (5). Continuarea gluconeogenezei pentru formarea metaboliților derivați din hexoză este un proces important ingerminarea semințelor (Buchanan și colab., 2015; Eastmond & Graham, 2001; Quettier & Eastmond, 2009):

catabolismul Fosfoenolpiruvatimplică o fosforilare la nivel de substrat care produce piruvat (etapa E-1,Figura 1), decarboxilarea oxidativă a piruvatului (etapa F-1) cu 1 Nadhproducție și oxidarea acetil-CoA la CO2. În oxidarea acetil-CoA, producția de 3 NADH, 1 FADH2 și 1 GTP corespunde la 10 fosfați de mare energie. Această valoare adăugată la nivelul substratului fosforilarea și fosforilarea oxidativă asociată cu decarboxilarea generată de NADH a piruvatului, totalizând 13,5 ATP (ecuația 6).

înlocuirea ecuațiilor (2), (3), (4),(5) și (6) în ecuația (1), se obține ecuația (7). Când este simplificatalgebric, ecuația (8) stabilește un raport direct între cantitatea de energie produsă (x) și dimensiunea lanțului de acizi grași (n).

validarea ecuației (8) pentru a calcula energia (ATP) produsă de acizii grașidegradarea prin ciclul glioxilat
astfel încât ecuația (8) să poată fi validată,cantitatea de ATP, obținută din aplicarea sa, a fost comparată cu cantitatea de ATP calculată individual din suma moleculelor acetil-CoA, NAHD și FADH2 și fosforilarea nivelului substratului, generată în oxidarea acizilor grași cu un număr specific de atomi de carbon în glioxizomi (Tabelul 2). Înambele condiții, sumele ATP obținute sunt echivalente. Luând în considerare aceste rezultate echivalente, ecuația dezvoltată în studiul actual a determinat cantitatea de energie produsă în catabolismul lipidic, având în vedere integrarea căilor metabolice care implică mai mult de douăzeci de pași.

mai mult, ecuațiile de echilibrare energetică, urmate de validarea lor, pot fi un instrument important pentru analiza diferitelor căi metabolice. În biochimie, aspecte cum ar fi relațiile stoichiometrice în căile metabolice pot fi abordate folosind rezultatele cercetărilor actuale.

Comparațieîntre eficiența degradării acizilor grași prin ciclul glioxilat și satabolismul animalic
conform studiilor anterioare, cantitatea de energie din celulele animale produsă în ATP prin degradarea acizilor grași cu numărul perechilor de atomi de carbon poate fi legată algebric de dimensiunea lanțului acidului gras. Această relație este exprimată prin ecuația (9) (Gonximalves și colab.,2012), care poate fi comparată cu ecuația (8) dezvoltată în studiul actual. Cele două ecuații pot fi utilizate pentru a compara eficiența energetică între oxidarea cu oxid de oxid în celulele animale și celulele organismelor, unde are loc ciclul glioxilat.

prin împărțirea ecuației (8) la ecuația(9) și înmulțirea cu 100, se obține procentul de eficiență (e%) a degradării acizilor grași pentru a produce energie, în raport cu catabolismul animal, așa cum este exprimat prin ecuația (10).

trasarea e % împotriva lungimii lanțului de acizi grași (Figura 2) verifică dacă eficiența scade odată cu creșterea lanțului de hidrocarburi. Mai mult, energia obținută din oxidarea acizilor grași în celulele cu ciclu glioxilat atinge mai puțin de 90% din nivelul de energie care ar fi generat de calea similară într-o celulă animală.
rata ATP produsă per atom de carbon oxidat (ATP/n) este, de asemenea, dependentă de lungimea lanțului acidului gras, la o rată maximă apropiată de șase ATP/n (Figura 3-A), în timp ce această rată este mai mare în celulele animale (Figura 3-b).

Figura 2.
comparatie intre eficienta energetica a degradarii acizilor grasi la animale-oxidarea si oxidarea la organisme cu enzime ale ciclului glioxilat, datorita lantului lung al acizilor grasi.
în contrast cu animalele, legumeleși unele microorganisme sunt capabile să transforme acetil-CoA din oxidarea acizilor grași în zaharuri. Organismele combină ciclul glioxilat și reacțiile de gluconeogeneză, care sunt compartimentate între glioxizomi / peroxizomi,mitocondrii și citosol. Această caracteristică oferă o versatilitate metabolică mai mare,permițând bacteriilor să utilizeze acetat ca sursă de energie și plante pentru a stoca lipide în semințe ca sursă de energie pentru a fi utilizate în timpul germinării.

Figura 3.
rata ATP produsă de atomul de carbon (ATP/n)datorită atomilor de carbon ai acizilor grași (n) în ciclul glioxilat (a) și catabolismul animalelor (b). Rata ATP / n crește semnificativ până laaproximativ n = 20, de acum înainte rata crește într-un mod mai puțin semnificativ.
concluzie
degradarea acizilor grași de către ciclul glioxilat pentru producerea de energie implică oxidarea cu oxidativ, cu caracteristicile sale particulare în glioxizomi, ciclul glioxilat, ciclul Krebs, gluconeogeneza și glicoliza. În ciuda acestor etape și căi metabolice, o relație stoichiometrică între lungimea lanțului acizilor grași și cantitatea de energie produsă în degradarea sa poate fi stabilită în termeni algebrici.
comparația dintre energetica stoichiometrică implicată în aceste căi metabolice și energetica stoichiometrică a oxidării cu oxigen așa cum apare în celulele animale, arată o reducere a eficienței energetice, în comparație cu metabolismul animal. În timp ce ocolirea reacțiilor de decarboxilare ale ciclului Krebs conferă adaptări evolutive organismelor atunci când sunt prezente, determină, de asemenea, o cantitate mai mică de energie generată pe atom de carbon în comparație cu degradarea acizilor grași la animale.